
Axít ribonucleic (ARN hay RNA) là một phân tử polymer cơ bản có nhiều vai trò sinh học trong mã hóa, dịch mã, điều hòa và biểu hiện gen. RNA và DNA đều là các axit nucleic, và cùng với lipid, protein và carbohydrat, tạo thành bốn loại phân tử cơ bản cho mọi dạng sự sống trên Trái Đất. Như DNA, RNA được tạo thành từ một chuỗi nucleotide, nhưng khác với DNA, RNA thường tự gấp lại thành một sợi đơn thay vì sợi kép. Các sinh vật sử dụng mRNA để truyền tải thông tin gen (sử dụng các nucleobase guanin, uracil, adenin và cytosin, được biểu thị bằng các chữ cái G, U, A và C) để tổng hợp protein trực tiếp. Nhiều loại virus lưu trữ thông tin gen của họ dưới dạng RNA gen.
Một số phân tử RNA hoạt động bên trong tế bào như là chất xúc tác sinh học, điều khiển biểu hiện gen hoặc tham gia vào các quá trình phản ứng và truyền tín hiệu tế bào. Một trong những chức năng chính là tổng hợp protein, mà RNA trực tiếp tham gia vào quá trình này trên ribosome. Quá trình này sử dụng tRNA để vận chuyển các axit amin đến ribosome, nơi mà rRNA ghép nối các axit amin lại với nhau để tạo thành chuỗi protein tiền.
So sánh với DNA
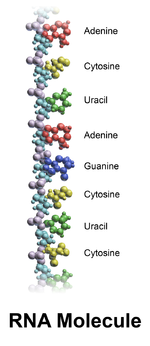
Cấu trúc hóa học của RNA có những điểm tương tự với DNA, nhưng có ba điểm khác biệt cơ bản:
- Không giống với sợi kép của DNA, RNA là phân tử sợi đơn trong hầu hết các chức năng sinh học và chứa chuỗi nucleotide ngắn hơn nhiều. Tuy nhiên, RNA có thể tạo thành cấu trúc sợi kép tự gập từ một sợi đơn bằng cách bổ sung các nền tảng tương ứng, như trong trường hợp của tRNA.
- Trong khi 'bộ khung' đường-phosphate của DNA chứa deoxyribose, bộ khung của RNA là phân tử ribose. Đường ribose có một nhóm hydroxyl gắn với vòng pentose ở vị trí 2', trong khi đối với phân tử deoxyribose thì không có. Nhóm hydroxyl trong đường khung ribose làm cho RNA ít ổn định hơn so với DNA do dễ bị thủy phân hơn.
- Nền tảng bổ sung adenine trong DNA là thymine, trong RNA là uracil, là dạng chưa methyl hóa của thymine.
Tương tự như DNA, hầu hết các hoạt động sinh học của RNA như mRNA, tRNA, rRNA, snRNA và các RNA không mã hóa khác, chứa các chuỗi bổ sung cho phép RNA tự gập lại và tạo thành sợi kép xoắn ốc. Phân tích các RNA này cho thấy chúng có cấu trúc bậc cao. Không giống với DNA, chúng không chứa một sợi kép quá dài, mà là một hệ bao gồm các sợi kép ngắn kết hợp với các cấu trúc tương tự như protein.
Theo cấu trúc này, RNA có thể đóng vai trò như các chất xúc tác (như enzyme). Ví dụ, khi xác định cấu trúc của ribosome — một phức hợp RNA-protein tham gia vào quá trình hình thành chuỗi peptide — các nhà sinh học đã phát hiện vị trí hoạt động của nó được hoàn toàn kiểm soát bởi RNA.
Cấu trúc hình thái
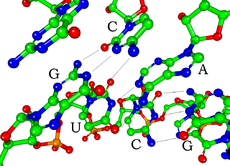
Mỗi nucleotide trong RNA chứa một đường ribose, với cacbon được đánh số từ 1' đến 5'. Nhìn chung, một base được gắn vào vị trí 1' là adenine (A), cytosine (C), guanine (G), hoặc uracil (U). Adenine và guanine là purine, cytosine và uracil là pyrimidine. Một nhóm phosphat gắn vào vị trí 3' của một đường ribose và vị trí 5' của đường ribose tiếp theo. Nhóm phosphat mang điện âm, khiến cho RNA là phân tử polyanion. Các base tạo thành liên kết hydrogen giữa cytosine và guanine, adenine và uracil, và guanine và uracil. Tuy nhiên, cũng có thể có các tương tác khác, như adenine liên kết với một nhóm khác tạo thành một vòng bốn (tetraloop) GNRA với cặp base guanine–adenine.
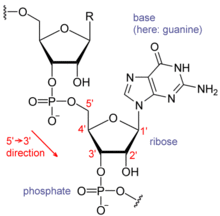
Một đặc điểm cấu trúc quan trọng của RNA khác với DNA là sự hiện diện của nhóm hydroxyl tại vị trí 2' trong đường ribose. Đặc tính này làm cho hình dạng A-hình học của RNA, mặc dù trong trường hợp của đơn dinucleotide, có thể hiếm gặp hình dạng B-hình học như quan sát trong DNA. Hình dạng A-hình học tạo ra rãnh lớn và sâu cũng như một rãnh nhỏ rộng và nông trên phân tử RNA. Sự hiện diện của nhóm 2'-hydroxyl cũng dẫn đến khả năng hóa học của các vùng linh hoạt về cấu trúc của RNA, có thể tấn công liên kết phosphodiester kế bên để cắt bỏ khung RNA.
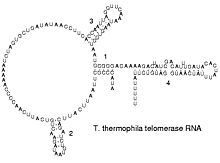
RNA được phiên mã chỉ từ bốn nucleotide (adenine, cytosine, guanine và uracil), nhưng cả base và nhóm ribose có thể được chỉnh sửa nhiều cách khi RNA trưởng thành. Ở pseudouridine (Ψ), mối liên kết uracil và ribose chuyển từ liên kết C–N thành liên kết C–C, và ribothymidine (T) được tìm thấy ở nhiều nơi (đặc biệt là ở vòng TΨC của tRNA). Một ví dụ khác về sự biến đổi base là hypoxanthine, một dạng khử amin của adenine tạo thành nucleoside inosine (I). Inosine đóng vai trò quan trọng trong giả thuyết cặp base linh hoạt (wobble hypothesis) trong mã di truyền.
Có hơn 100 dạng nucleoside biến đổi tồn tại trong tự nhiên. Sự đa dạng lớn nhất trong cấu trúc sửa đổi này thường được tìm thấy ở tRNA, trong khi pseudouridine và nucleoside với 2'-O-methylribose phổ biến nhất trong rRNA. Các nhà sinh học vẫn chưa hoàn toàn hiểu vai trò đặc thù của nhiều biến đổi này trong RNA. Tuy nhiên, đáng chú ý là, trong RNA ribosome, nhiều thay đổi sau khi phiên mã xảy ra ở những vùng có chức năng quan trọng như trung tâm peptidyl transferase và giao diện tiểu đơn vị, ngụ ý rằng chúng quan trọng đối với chức năng bình thường.
Cấu trúc chức năng của các phân tử RNA dạng sợi đơn, tương tự như protein, thường yêu cầu một cấu trúc bậc ba cụ thể. Các khung cho cấu trúc này được cung cấp bởi các yếu tố cấu trúc bậc hai như liên kết hydro trong phân tử. Điều này dẫn đến sự xuất hiện của một số 'miền' có thể nhận biết được của cấu trúc bậc hai như vòng kẹp tóc, phình và vòng lặp nội bộ. Vì RNA mang điện tích, sự hiện diện của ion kim loại như Mg là cần thiết để ổn định nhiều cấu trúc bậc hai và bậc ba của RNA.
Dạng đồng phân lập thể enantiomer tự nhiên của RNA là D-RNA, trong đó các D-ribonucleotide được sử dụng. Tất cả các trung tâm đối xứng nằm trong D-ribose. Bằng cách sử dụng L-ribose hoặc L-ribonucleotide, có thể tổng hợp được L-RNA. L-RNA có độ ổn định cao hơn đối với sự phân hủy của RNase.
Như những phân tử sinh học khác như protein, cấu trúc của RNA có thể được xác định bởi mạch tô pô của nó khi đã gấp. Điều này thường dựa trên sự sắp xếp các vị trí tiếp xúc nội chuỗi bên trong RNA đã gấp, được gọi là topology mạch tô pô.
Quá trình tổng hợp
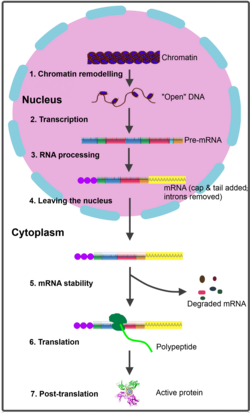
Phiên mã là quá trình tổng hợp RNA, nơi enzym polymerase RNA sử dụng một mẫu gen trên DNA làm mẫu. Quá trình bắt đầu với enzym kết nối vào trình khởi động trên DNA ở phía 'thượng nguồn' của gen.
- Để bắt đầu phiên mã, chuỗi xoắn kép của DNA ở vùng gen cần phiên mã phải được tháo xoắn bởi topoisomerase, sau đó được dãn mạch và tách đôi bởi enzym helicase.
- Enzym polymerase RNA di chuyển dọc theo mạch mẫu (mạch gốc) theo chiều từ 3’ đến 5’ của gen, tổng hợp chuỗi polyribonucleotide theo nguyên tắc bổ sung, kéo dài theo hướng từ 5’ đến 3’ (ngược lại với hướng di chuyển của nó). Thứ tự của các deoxyribonucleotide trên mạch gốc của gen không chỉ quyết định thứ tự chuỗi polyribonucleotide của RNA mà còn quy định cả sự kết thúc của quá trình phiên mã.
- Ở tế bào nhân, RNA mới được tổng hợp gọi là pre-RNA hoặc RNA sơ khai. Nó cần trải qua quá trình gọi là sửa đổi sau khi tổng hợp để tạo thành RNA trưởng thành. Trong quá trình xử lý RNA:
- Đầu 5' của nó được gắn với GTP để tạo 'mũ', còn đầu 3' của nó được gắn với pôlyA để tạo 'đuôi'.
- RNA sơ khai phải bị loại bỏ các intron (vùng không mã hóa), và các intron này sẽ được giải phân.
- Các intron (đoạn không mã hóa) của nó bị cắt bỏ, trong khi các exon (đoạn có mã hóa) được nối lại với nhau để tạo thành một chuỗi bộ ba mã di truyền liên tục. Giai đoạn này được gọi là cắt nối RNA.
Quá trình này phụ thuộc vào nhiều nhân tố, nhưng quan trọng nhất là spliceosome (một tổ hợp phân tử lớn và phức tạp). Sau khi hoàn tất quá trình sửa đổi, RNA trưởng thành được hình thành và được xuất ra ngoài tế bào qua hố nhân.
Trong một số nhóm sinh vật ít, có một số enzyme RNA phụ thuộc RNA (RNA-dependent RNA polymerase) sử dụng RNA làm mẫu để tổng hợp RNA mới. Ví dụ, một số virus RNA (như poliovirus) sử dụng enzyme này để sao chép vật liệu di truyền của chúng. Tương tự, RNA polymerase phụ thuộc RNA là một phần quan trọng của quá trình can thiệp RNA ở nhiều sinh vật.
Các dạng RNA
Tổng quát về RNA
RNA tin nhắn (mRNA) là loại RNA mang thông tin từ DNA đến ribosome, nơi diễn ra quá trình dịch mã để tổng hợp protein trong tế bào. Trình tự mã hóa của mRNA xác định thứ tự amino acid trong protein được tổng hợp. Tuy nhiên, nhiều RNA không phải mã hóa protein (khoảng 97% sản phẩm RNA từ quá trình biểu hiện gen là các protein không mã hóa trong sinh vật nhân thực).
Các RNA không mã hóa (ncRNA) có thể được mã hóa bởi gene RNA của chúng hoặc được tạo thành từ các intron mRNA. Ví dụ điển hình cho các RNA không mã hóa là tRNA và rRNA, cả hai tham gia vào quá trình dịch mã. Các RNA không mã hóa cũng có vai trò trong điều hòa biểu hiện gen, xử lý RNA và các hoạt động khác. Một số RNA còn có khả năng hoạt động như enzyme, tham gia vào các phản ứng sinh học như cắt và nối các phân tử RNA khác, cũng như tham gia vào quá trình tạo liên kết peptide trong ribosome, được biết đến với tên gọi ribozyme.
Theo chiều dài
RNA có thể được phân thành RNA nhỏ và RNA dài dựa trên độ dài của chúng. RNA nhỏ thường có độ dài dưới 200 nt, trong khi RNA dài có độ dài hơn 200 nt. RNA dài bao gồm chủ yếu lncRNA và mRNA. RNA nhỏ bao gồm các loại như rRNA 5.8S, 5S rRNA, tRNA, miRNA, siRNA, snoRNAs, piRNA, tsRNA và srRNA.
Theo quá trình dịch mã

mRNA chuyển thông tin về trình tự protein tới ribosome để tổng hợp protein trong tế bào. Mỗi ba nucleotide tương ứng với một amino acid. Trong sinh vật nhân thực, pre-mRNA được phiên mã từ DNA, sau đó xử lý để trở thành mRNA trưởng thành bằng cách loại bỏ intron. mRNA sau đó được vận chuyển từ nhân tế bào ra bào tương, nơi nó gặp ribosome để dịch mã thành protein với sự tham gia của tRNA. Trong sinh vật nhân sơ, mRNA có thể gắn trực tiếp vào ribosome trong khi vẫn được phiên mã từ DNA. Sau một thời gian, mRNA này sẽ bị phân hủy thành các nucleotide với sự trợ giúp của ribonuclease.
tRNA là RNA nhỏ khoảng 80 nucleotide vận chuyển amino acid đến ribosome để tham gia vào tổng hợp protein trong quá trình dịch mã. Nó có vùng gắn amino acid và anticodon nhận diện codon trên mRNA thông qua liên kết hydro.
rRNA là thành phần xúc tác của ribosome. Trong sinh vật nhân thực, ribosome chứa các phân tử rRNA như 18S, 5.8S, 28S và 5S. Ba phân tử rRNA được tổng hợp trong nhân con và phân tử còn lại được tổng hợp ở nơi khác. Trong bào tương, rRNA và protein kết hợp lại thành ribosome, gắn với mRNA và thực hiện tổng hợp protein.
tmRNA được tìm thấy ở nhiều vi khuẩn và lạp thể. Nó đánh dấu các protein do mRNA mã hóa mà thiếu codon kết thúc, ngăn cản ribosome bị dừng sớm.
RNA điều hòa
Một số loại RNA có khả năng điều hòa quá trình biểu hiện gen bằng cách liên kết bổ sung với một phần của mRNA hoặc DNA gen. MicroRNA (miRNA; dài 21-22 nt) đã được phát hiện ở sinh vật nhân thực và hoạt động thông qua can thiệp RNA (RNAi), nơi mà một hệ thống phức hợp của miRNA và các enzyme có thể cắt mRNA, ngăn chặn quá trình dịch mã của mRNA hoặc tăng tốc độ phân hủy của nó.
Trong khi RNA can thiệp nhỏ (small interfering RNA, siRNA; 20-25 nt) thường được tạo ra bằng cách phá RNA của virus, cũng có nguồn nội sinh siRNA. siRNA hoạt động bằng cách can thiệp RNA giống như miRNA. Một số miRNA và siRNA có thể ảnh hưởng đến sự methyl hóa của gen, dẫn đến sự giảm hoặc tăng hoạt động biên dịch của các gen này. Ở động vật có RNA tương tác Piwi (piRNA; 29-30 nt), chúng hoạt động trong tế bào gốc (germline) và có vai trò quan trọng trong việc chống lại transposon và trong quá trình hình thành gamete.
Nhiều sinh vật nhân sơ có hệ thống RNA CRISPR, một hệ thống điều hòa tương tự như can thiệp RNA. RNA đối nghịch (antisense RNA) được tìm thấy rộng rãi, trong đó hầu hết làm giảm hoạt động của gen nhưng cũng có một số tăng hoạt động phiên mã. Một cách RNA đối nghịch hoạt động khi nó gắn vào mRNA, tạo thành một sợi RNA kép giảm chức năng của enzyme. Có nhiều loại RNA không mã hóa dài tham gia điều hòa gen ở sinh vật nhân thực, ví dụ như RNA Xist, làm lạnh nhiễm sắc thể X ở cái và góp phần vào việc bất hoạt nó.
Một mRNA có thể chứa các phần tử điều hòa trong chính nó, như các riboswitch nằm ở vùng không dịch mã ở đầu 5' hoặc 3' của mRNA; các yếu tố điều hòa trong vùng này điều khiển hoạt động của mRNA. Các vùng không dịch mã cũng có thể chứa các phần tử tham gia điều hòa gen ở các gen khác.
Theo quá trình xử lý RNA
Nhiều loại RNA tham gia vào quá trình sửa đổi của các RNA khác. Các đoạn intron được cắt ra từ pre-mRNA bởi spliceosome, trong đó có mặt các RNA hạt nhân nhỏ (snRNA), hoặc các intron có thể là ribozyme được sử dụng để cắt các đoạn intron khác. RNA cũng có thể được chỉnh sửa bằng cách thay đổi các nucleotide A, C, G và U từ một RNA này sang một RNA khác. Ở sinh vật nhân thực, quá trình chỉnh sửa nucleotide của RNA được điều khiển chủ yếu bởi các RNA nucleolar nhỏ (snoRNA; 60–300 nt), được tìm thấy trong nhân con và các thể Cajal. snoRNA phối hợp với các enzyme đến một vị trí trên RNA bằng cách ghép cặp cơ sở với RNA. Các enzyme này sau đó thực hiện sửa đổi nucleotide. rRNA và tRNA là những phân tử phổ biến bị sửa đổi, và snRNA cũng có thể là mục tiêu của sửa đổi base.
Bộ gene của RNA
Giống như DNA, RNA có thể chứa thông tin di truyền. Các virus RNA có bộ gene chứa RNA mã hóa cho các protein của chúng. Bộ gene của virus được sao chép bằng một số protein, trong khi các protein khác có vai trò bảo vệ bộ gene khi hạt virus chuyển sang tế bào chủ mới. Viroid là một nhóm tác nhân gây bệnh khác, chỉ chứa RNA mà không mã hóa bất kỳ protein nào, và được sao chép bởi polymerase của tế bào thực vật chủ.
Theo quá trình transkripsi ngược
Các virus chứa bộ gene RNA phải trải qua quá trình tổng hợp ngược thành DNA bổ sung bởi enzyme phiên mã ngược; sau đó, DNA này được phiên mã thành RNA mới để thực hiện quá trình dịch mã. Retrotransposon lan rộng thông qua sao chép DNA và RNA từ một tế bào sang tế bào khác, trong khi telomerase chứa một RNA dùng làm mẫu để tái tạo các đoạn cuối của nhiễm sắc thể ở sinh vật nhân thực.
RNA đôi sợi
RNA đôi sợi (dsRNA) là RNA có hai sợi bổ sung, tương tự như DNA trong mọi tế bào. dsRNA là vật liệu di truyền chính ở một số virus (virus RNA đôi sợi, double-stranded RNA viruses). RNA đôi sợi như trong virus RNA hoặc siRNA có thể kích hoạt can thiệp RNA ở sinh vật nhân thực, cũng như kích hoạt các protein interferon trong động vật có xương sống.
RNA vòng
Vào cuối những năm 1990, các nhà sinh học đã phát hiện một loại RNA đơn sợi khép kín ở động vật. Sau đó, loại RNA này đã được chính thức xác định và được gọi là RNA vòng (circRNA). Chi tiết xem tại trang RNA vòng.
Những khám phá quan trọng về RNA sinh học

Nghiên cứu về RNA đã dẫn đến nhiều khám phá sinh học quan trọng và cũng nhiều giải Nobel. Acid nucleic được Friedrich Miescher khám phá lần đầu tiên vào năm 1868, ông gọi chúng là 'nuclein' do được tìm thấy trong nhân tế bào. Sau đó phát hiện ra rằng trong sinh vật nhân sơ, không có nhân cũng có chứa acid nucleic. Giả thuyết về vai trò của RNA trong quá trình tổng hợp protein đã được nêu ra từ năm 1939. Severo Ochoa nhận giải Nobel Sinh lý học và Y học năm 1959 (cùng với Arthur Kornberg) cho khám phá về enzyme cho phép tổng hợp RNA trong phòng thí nghiệm. Tuy nhiên, loại enzyme được Ochoa khám phá (polynucleotide phosphorylase) sau này đã được chứng minh có vai trò làm thoái hóa RNA, không phải tổng hợp RNA. Năm 1956, Alex Rich và David Davies cho lai hai dòng RNA để tạo thành tinh thể RNA đầu tiên, cấu trúc của nó có thể xác định bằng kỹ thuật nhiễu xạ tia X (tinh thể học tia X).
Trình tự 77 nucleotide trong tRNA của một loài nấm men được Robert W. Holley xác định lần đầu vào năm 1965, giúp ông đoạt giải Nobel Sinh lý học và Y học năm 1968 (cùng với Har Gobind Khorana và Marshall Nirenberg).
Vào đầu những năm 1970, retrovirus và enzyme phiên mã ngược được phát hiện, lần đầu tiên chứng minh rằng enzyme tham gia quá trình sao chép từ RNA sang DNA (quá trình ngược lại so với chu trình thông thường của truyền thông tin di truyền). Nhờ vào khám phá này, David Baltimore, Renato Dulbecco và Howard Temin đã đoạt giải Nobel Y học năm 1975.
Năm 1976, Walter Fiers và các nhà khoa học đã thành công trong việc giải trình tự RNA trong một gene của virus, cụ thể là bacteriophage MS2.
Năm 1977, các intron và quá trình ghép RNA (RNA splicing) được phát hiện ở cả virus đối với động vật và ở gene tế bào, mang đến giải Nobel cho Philip Sharp và Richard Roberts vào năm 1993. Các phân tử RNA xúc tác (ribozyme) được phát hiện đầu những năm 1980, và đã giúp Thomas Cech và Sidney Altman đoạt giải Nobel năm 1989. Năm 1990, một khám phá quan trọng là trong cây Petunia (dã yên thảo) cho thấy có thể dùng các gene để tắt các gene tương tự trong cùng loài thực vật này, mở đường cho kỹ thuật can thiệp RNA sau này.
Trong cùng thời kỳ này, các RNA nhỏ 22 nucleotide, hiện được biết đến như microRNA, được phát hiện có vai trò quan trọng trong sự phát triển của C. elegans.
Nghiên cứu về can thiệp RNA đã dẫn tới giải Nobel Y học năm 2006 cho Andrew Fire và Craig Mello, và giải Nobel Hóa học cho Roger Kornberg cùng năm. Khám phá về các RNA điều hòa biểu hiện gene đã thúc đẩy sự phát triển của các loại thuốc từ RNA, như siRNA, có chức năng làm tắt một số gene.
Liên quan đến hóa học tiền sinh học và thuyết phát sinh sinh vật
Năm 1967, Carl Woese đưa ra giả thuyết rằng RNA có thể là chất xúc tác và gợi ý về sự tồn tại của các sinh vật nguyên thủy nhất dựa trên RNA, không chỉ chứa thông tin di truyền mà còn là chất xúc tác cho các phản ứng hóa sinh — được gọi là giả thuyết thế giới RNA.
Tháng 3 năm 2015, các nucleotide phức tạp của DNA và RNA, bao gồm uracil, cytosine và thymine, được thông báo đã tổng hợp thành công trong phòng thí nghiệm dưới điều kiện của không gian ngoài trái đất, sử dụng các hợp chất hữu cơ ban đầu như pyrimidine, một hợp chất phổ biến được tìm thấy trong các vật liệu hữu cơ như vật liệu có nhiều vòng naphthalene (PAHs). Pyrimidine, giống như các hydrocarbon thơm đa vòng (PAHs), là một trong những hợp chất giàu carbon nhất được phát hiện trong Vũ trụ và có thể hình thành trong môi trường xung quanh các ngôi sao đỏ hoặc các đám mây khí bụi giữa các ngôi sao.
- Cấu trúc sinh học của RNA
- Origami RNA
- Đại phân tử
- DNA
- Transcriptome
Liên kết ngoài
- RNA tại Từ điển bách khoa Việt Nam
- RNA (sinh hóa) tại Encyclopædia Britannica (bằng tiếng Anh)
- Trang web Thế giới RNA Bộ sưu tập liên kết (cấu trúc, chuỗi, công cụ, tạp chí)
- Cơ sở dữ liệu Acid nucleic Hình ảnh về DNA, RNA và các phức tạp.
- Seminar của Anna Marie Pyle: Cấu trúc, chức năng và nhận diện RNA
Chủng loại Acid nucleic |
|---|
Di truyền học |
|---|
Biểu hiện gen |
|---|
Cấu trúc của tế bào / bào quan |
|---|
Kháng sinh: chất ức chế axit nucleic (J01E, J01M) |
|---|


Sinh học phân tử |
|---|
| Tiêu đề chuẩn |
|
|---|