

| Bạc má lớn | |
|---|---|
| Con mái ở Lancashire, Anh. | |
Tình trạng bảo tồn
| |
Ít quan tâm (IUCN 3.1) | |
| Phân loại khoa học | |
| Vực: | Eukaryota |
| Giới: | Animalia |
| Ngành: | Chordata |
| Lớp: | Aves |
| Bộ: | Passeriformes |
| Họ: | Paridae |
| Chi: | Parus |
| Loài: | P. major
|
| Danh pháp hai phần | |
| Parus major Linnaeus, 1758 | |
| Phạm vi phân bố các phân loài trước đây và hiện tại của bạc má lớn | |
Bạc má lớn (danh pháp hai phần: Parus major) là một loài chim trong họ Bạc má. Bạc má lớn là một loài phổ biến ở châu Âu, Trung Đông, Trung Á, phân bố đến tận phía đông khu vực sinh thái Cổ Bắc giới, sông Amur và phân bố tại một phần Bắc Phi. Bạc má lớn hầu như không di trú nếu mùa đông không quá khắc nghiệt. Trước năm 2005, loài này được gộp chung với rất nhiều phân loài khác. Các nghiên cứu DNA cho thấy sự khác biệt giữa một số phân loài của bạc má lớn. Hiện nay các phân loài này đã được tách ra thành hai loài riêng biệt là bạc má lưng xám (Parus cinereus) ở Nam Á và bạc má Nhật Bản (Parus minor) ở Đông Á. Bạc má lớn vẫn là loài phổ biến nhất trong chi Parus.
Bạc má lớn là loài chim có đặc điểm: đầu đen, cổ và má trắng, lưng màu ô liu và thân dưới màu vàng, có một số khác biệt giữa các phân loài. Loài chim này chủ yếu ăn côn trùng vào mùa hè, ăn nhiều loại thức ăn hơn trong những tháng mùa đông, chim ăn cả con dơi nhỏ đang ngủ đông. Giống như các loài bạc má khác, bạc má lớn làm tổ trong hốc cây. Chim mẹ đẻ khoảng 12 quả trứng và ấp trứng một mình. Cả chim bố và chim mẹ đều nuôi chim non. Mỗi năm chim sinh hai lứa. Tổ của chúng có thể bị gõ kiến nhỏ sườn đỏ, sóc xám miền Đông và triết bụng trắng tấn công, và bị bọ chét ký sinh. Chim trưởng thành bị chim cắt hỏa mai săn bắt. Bạc má lớn đã thích nghi tốt với những thay đổi của con người đối với môi trường, và là loài chim phổ biến và quen thuộc trong các công viên và khu vườn đô thị. Bạc má lớn cũng là một loài nghiên cứu quan trọng trong ngành điểu học.
Phân loại

Theo phiên bản thứ 10 của Systema Naturae năm 1758, Carl Linnaeus đặt danh pháp khoa học cho bạc má lớn là Parus major. Tên khoa học của loài có nguồn gốc từ tiếng Latinh: parus (bạc má) và maior (rộng lớn). Francis Willughby đã sử dụng tên này vào thế kỷ 17.
Bạc má lớn trước đây được cho là phân bố từ Anh đến Nhật Bản và phía nam đến các đảo của Indonesia, với 36 phân loài được mô tả thuộc bốn nhóm phân loài chính. Nhóm major có 13 phân loài phân bố trên khắp châu Âu, vùng ôn đới châu Á và Bắc Phi. Nhóm minor với 9 phân loài phân bố từ đông nam Nga và Nhật Bản đến phía bắc của đông nam châu Á. Nhóm cinereus với 11 phân loài được tìm thấy từ Iran qua Nam Á đến Indonesia. Ba phân loài bokharensis thường được coi là một loài riêng biệt, Parus bokharensis. Cách phân loại này từng được cho là tạo thành một loài vành đai quanh Cao nguyên Tây Tạng, với dòng gen chung xuyên suốt các phân loài. Tuy nhiên, lý thuyết này đã bị bác bỏ khi việc kiểm tra các chuỗi DNA ty thể đã phát hiện ra rằng bốn nhóm là đơn ngành, và các vùng lai giữa các nhóm là kết quả của quá trình lai thứ cấp sau khoảng thời gian tạm tách biệt nhau.

Một nghiên cứu được công bố vào năm 2005 đã xác nhận sự khác biệt giữa nhóm major với nhóm cinereus và nhóm minor, và cùng với phân loài P. m. bokharensis tách khỏi hai nhóm này vào khoảng 1,5 triệu năm trước. Sự khác biệt giữa nhóm bokharensis và các nhóm khác được ước tính là từ khoảng nửa triệu năm trước. Nghiên cứu cũng kiểm tra các giống lai của đại diện các nhóm major và minor ở Thung lũng Amur, nơi có sự giao thoa của hai nhóm. Kết quả cho thấy con lai rất hiếm, tức là có một số rào cản sinh sản giữa hai nhóm. Nghiên cứu khuyến nghị tách hai nhóm phía đông thành loài mới bao gồm: bạc má lưng xám (Parus cinereus) và bạc má Nhật Bản (Parus minor). Riêng bạc má Turkestan thì vẫn gộp chung với bạc má lớn. Cách phân loại này đã được một số cơ quan có thẩm quyền tuân theo, như là Danh sách các loài chim trên thế giới của IOC (IOC World Bird List). Cẩm nang về các loài chim trên thế giới (Handbook of the Birds of the World) thì phân loại các loài thuộc chi Parus theo cách ban đầu: coi bạc má Turkestan là một loài riêng biệt, nhưng vẫn gộp bạc má Nhật Bản và bạc má lưng xám với bạc má lớn, các phân loại này gây nhiều tranh cãi.
Phân loài đại diện của bạc má lớn là phân loài phổ biến nhất, với phạm vi trải dài từ bán đảo Iberia đến thung lũng Amur và từ Scandinavia đến Trung Đông. Các phân loài khác phân bố hẹp hơn nhiều, bốn phân loài có phân bố hạn chế ở các đảo và phần còn lại của phân loài P. m. major phân bố ở các vùng biệt cư để tránh kỷ băng hà trước đây. Sự phổ biến của một phân loài duy nhất, đồng nhất về mặt hình thái trên một khu vực rộng lớn như vậy cho thấy rằng phân loài đại diện đã nhanh chóng tái định cư trên một khu vực rộng lớn sau kỷ băng hà cuối cùng. Giả thuyết này được hỗ trợ bởi các nghiên cứu di truyền cho thấy một thắt cổ chai di truyền gần đây về mặt địa chất, dẫn đến sự gia tăng dân số nhanh chóng.
Chi Parus từng bao gồm hầu hết các loài chim bạc má trong họ Paridae, nhưng các nghiên cứu về hình thái và di truyền đã dẫn đến việc chia tách chi lớn vào năm 1998. Bạc má lớn được giữ lại trong Parus và cùng với Cyanistes tạo thành một dòng chim được biết đến là 'những loài không tích trữ', có liên quan đến tập tính tích trữ của các loài ở nhánh khác. Chi Parus vẫn là chi lớn nhất trong họ, nhưng có thể tiếp tục bị tách ra. Nếu không tính những loài trước đây được coi là phân loài, họ hàng gần nhất của bạc má lớn là bạc má cánh trắng (Machlolophus nuchalis) và bạc má lưng xanh (Parus monticolus) ở miền nam châu Á. Các giống lai với các loài ngoài chi Parus rất hiếm nhưng đã được ghi nhận với sẻ ngô xanh, sẻ ngô than và có thể là bạc má đầm lầy (Poecile palustris).
Phân loài

Hiện có 15 phân loài được công nhận:
- P. m. newtoni (Pražák, 1894), được tìm thấy khắp quần đảo Anh.
- P. m. major (Linnaeus, 1758), được tìm thấy ở khắp châu Âu, Tiểu Á, bắc và đông Kazakhstan, nam Siberia, bắc Mông Cổ cho đến tận giữa Thung lũng Amur.
- P. m. excelsus (Buvry, 1857), được tìm thấy ở tây bắc châu Phi.
- P. m. corsus (Kleinschmidt, 1903), được tìm thấy ở Bồ Đào Nha, phía nam Tây Ban Nha, và Corse.
- P. m. mallorcae (von Jordans, 1913), được tìm thấy ở quần đảo Baleares.
- P. m. ecki (von Jordans, 1970), được tìm thấy ở Sardegna.
- P. m. niethammeri (von Jordans, 1970), được tìm thấy ở Crete.
- P. m. aphrodite (Madarász, 1901), được tìm thấy ở phía nam Ý, phía nam Hy Lạp, Cộng hòa Síp và quần đảo Aegea.
- P. m. terrasanctae (Hartert, 1910), được tìm thấy ở Liban, Israel, Jordan và Syria.
- P. m. karelini (Zarudny, 1910), được tìm thấy ở đông nam Azerbaijan và tây bắc Iran.
- P. m. blandfordi (Pražák, 1894), được tìm thấy ở trung bắc trung và tây nam Iran.
- P. m. bokharensis (Lichtenstein, 1823), được tìm thấy ở nam Kazakhstan, Uzbekistan, Turkmenistan và viễn bắc Iran và Afghanistan. Cùng với hai phân loài dưới đây, cả ba từng được xem là một loài riêng.
- P. m. turkestanicus (Zarudny & Loudon, 1905), được tìm thấy từ đông Kazakhstan đến cực tây bắc Trung Quốc và tây Mông Cổ.
- P. m. ferghanensis (Buturlin, 1912), được tìm thấy ở Tajikistan và Kyrgyzstan.
- P. m. kapustini (Portenko, 1954), được tìm thấy từ tây bắc Trung Quốc (tây bắc Tân Cương) đến Mông Cổ và Siberia.
Mô tả

Bạc má lớn dài từ 12,5 đến 14,0 cm (4,9–5,5 in), và có vẻ ngoài đặc biệt giúp dễ dàng nhận ra. Phân loài đại diện P. m. major có đỉnh đầu màu xanh đen, cổ, họng, yếm và đầu màu đen, má và vành tai màu trắng. Ngực có màu vàng chanh sáng và có một sọc rộng màu đen ở giữa chạy từ yếm đến mông. Có một đốm trắng mờ trên cổ chuyển sang màu vàng xanh ở phía trên gáy. Phần còn lại của gáy và lưng có màu xanh ô liu. Phần trên của cánh có màu xanh lá cây, phần dưới của cánh có màu xám xanh với vạch cánh màu trắng. Đuôi có màu xám xanh với vành ngoài màu trắng. Bộ lông của con cái tương tự như của con đực, ngoại trừ màu sắc tổng thể nhạt hơn, yếm có màu đen nhạt hơn, cũng như sọc chạy xuống bụng cũng hẹp hơn và đôi khi bị đứt đoạn. Chim non giống chim mái, ngoại trừ việc chúng có gáy và cổ màu nâu ô liu xỉn, mông màu xám và đuôi xám hơn, với các đầu màu trắng không rõ ràng.

Có một số khác biệt giữa các phân loài. P. m. newtoni giống như phân loài đại diện nhưng có mỏ dài hơn một chút, phần trên cánh có màu xanh đậm hơn một chút, vành đuôi ít trắng hơn, và sọc giữa bụng rộng hơn. P. m. corsus cũng giống phân loài đại diện nhưng có phần trên xỉn màu hơn, vành đuôi ít trắng hơn và gáy ít vàng hơn. P. m. mallorcae giống phân loài đại diện, nhưng có mỏ lớn hơn, phần trên xanh xám hơn và phần dưới hơi nhạt hơn. P. m. ecki giống như P. m. mallorcae ngoại trừ phần trên xanh hơn và phần dưới nhạt hơn. P. m. excelsus tương tự như phân loài đại diện, nhưng có phần trên có màu xanh lá cây sáng hơn nhiều, phần dưới màu vàng sáng và không có (hoặc rất ít) màu trắng ở đuôi. P. m. aphrodite có phần trên sẫm màu hơn, xám ô liu hơn và phần dưới có màu vàng kem nhạt hơn. P. m. niethammeri tương tự như P. m. aphrodite nhưng phần trên xanh nhạt hơn, còn phần dưới màu vàng nhạt. P. m. terrasanctae giống với hai phân loài trước đó nhưng có phần trên hơi nhạt hơn. P. m. blandfordi giống như phân loài đại diện nhưng có phần trên cánh xám hơn, có phần ngang vai và phần dưới màu vàng nhạt, còn P. m. karelini là trung gian giữa phân loài đại diện và P. m. blandfordi, và không có vành trắng ở đuôi. P. m. bokharensis có bộ lông xám hơn nhiều, phần dưới có màu trắng kem nhạt đến xám, một mảng màu trắng lớn hơn, đuôi, cánh, lưng và gáy màu xám. Phân loài cũng nhỏ hơn một chút, mỏ nhỏ hơn nhưng đuôi dài hơn. Điều này cũng tương tự đối với hai phân loài có liên quan trong nhóm bạc mái Turkestan. P. m. turkestanicus giống P. m. bokharensis nhưng có mỏ lớn hơn và phần trên sẫm màu hơn. P. m. ferghanensis giống P. m. bokharensis nhưng có mỏ nhỏ hơn, màu xám đậm hơn ở hai bên sườn và màu vàng hơn ở những con chim non.

Màu ngực của chim trống đã được chứng minh là có tương quan với việc có tinh trùng khỏe mạnh hơn, và là một cách mà chim trống thể hiện ưu thế sinh sản của mình với chim mái. Hàm lượng carotenoid cao hơn làm tăng sắc vàng ở ngực, đồng thời giúp tinh trùng chống chọi tốt hơn với sự tấn công của các gốc tự do. Chim không thể tổng hợp carotenoid mà phải lấy từ thức ăn, vì vậy màu sắc càng tươi sáng ở chim trống chứng tỏ khả năng hấp thụ dinh dưỡng của chúng càng tốt. Tuy nhiên, độ bão hòa của màu vàng cũng bị ảnh hưởng bởi các yếu tố môi trường, chẳng hạn như điều kiện thời tiết. Chiều rộng sọc bụng của con đực, khác biệt ở từng cá thể, là một tiêu chí để con cái lựa chọn, với những con cái tốt hơn dường như chọn những con đực có sọc rộng hơn.
Tiếng hót

Bạc má lớn, giống như các loài bạc má khác, là loài chim biết hót, thậm chí có đến 40 kiểu kêu và giọng hót khác nhau. Tiếng kêu của chúng nói chung giống nhau giữa hai giới tính, tuy nhiên, con đực thường kêu nhiều hơn và con cái hiếm khi kêu. Các nốt đơn nhẹ nhàng như 'pit', 'spick' hoặc 'chit' thường được sử dụng để liên lạc. Tiếng 'tink' to hơn thường được con đực trưởng thành sử dụng để báo động hoặc trong các cuộc tranh chấp lãnh thổ. Một trong những tiếng kêu phổ biến nhất là 'teacher, teacher', thường được ví như tiếng bánh xe rùa, dùng để tuyên bố quyền sở hữu lãnh thổ. Ở Anh, người ta trước đây coi tiếng kêu 'mài cưa' là điềm báo trước mưa. Dù có sự khác biệt địa lý nhỏ, những con cá thể của hai nhóm Nam Á gần đây đã phân biệt riêng không nhận ra hoặc phản ứng với tiếng kêu của nhóm ôn đới.
Một giải thích cho sự phong phú của tiếng kêu của bạc má lớn là giả thuyết Beau Geste. Giả thuyết này được đặt tên từ tiểu thuyết phiêu lưu có tựa đề Beau Geste của nhà văn Anh P. C. Wren, trong đó nhân vật chính Michael 'Beau' Geste đã dựng xác những người lính đã ngã xuống để xây dựng các trận địa nhằm đánh lừa phe địch rằng pháo đài đang được bảo vệ tốt. Tương tự như vậy, sự đa dạng của tiếng hót cho thấy rằng lãnh thổ của bạc má có thể có mật độ cá thể dày đặc hơn so với thực tế. Dù giả thuyết này có đúng hay không, những con chim có khả năng hót nhiều sẽ chiếm ưu thế xã hội và thành công hơn trong sinh sản.
Phân bố, môi trường sống và di chuyển


Bạc má lớn phân bố rộng khắp lục địa Á-Âu. Chúng được tìm thấy trên khắp châu Âu trừ Iceland và phía bắc Scandinavia, bao gồm cả nhiều hòn đảo ở Địa Trung Hải. Tại Bắc Phi, loài được tìm thấy ở Maroc, Algérie và Tunisia. Nó cũng hiện diện ở Trung Đông, khu vực ở Trung Á từ phía bắc Iran và Afghanistan đến Mông Cổ, trên khắp miền bắc Á từ dãy núi Ural đến phía đông tại miền bắc Trung Quốc và Thung lũng Amur.

Bạc má lớn xuất hiện tại nhiều loại các môi trường sống. Loài này thường sống trong rừng thưa, rừng hỗn hợp, vùng ven rừng và các khu vườn. Chim cũng được tìm thấy ở những khu rừng dày đặc, bao gồm rừng tùng. Ở miền Bắc Siberia, loài được tìm thấy ở các cánh rừng taiga ở phía bắc. Tại Bắc Phi, loài có mặt ở rừng sồi cũng như là các khu vực tuyết tùng Atlas và thậm chí cả lùm cây cau. Ở phía đông bao gồm Siberia, Mông Cổ và Trung Quốc, chúng ưa thích rừng liễu và rừng bạch dương ven sông. Các phân loài Turkestan ở Trung Á cũng cư trú ở các cánh rừng này, cũng như cây bụi thấp, ốc đảo. Ở nơi cao hơn, chúng có mặt ở các môi trường sống khác nhau, từ rừng rụng lá rậm và rừng lá kim đến các khu vực mở có cây phân tán.
Bạc má lớn thường không di cư. Các cặp chim thường sẽ vẫn ở gần hoặc trong lãnh thổ của chúng quanh năm, ngay cả trong các khu vực phía bắc trong phạm vi phân bố của chúng. Chim non sẽ rời lãnh thổ của chim bố mẹ, nhưng thường không xa. Các quần thể bạc má lớn lên đến cả ngàn cá thể có thể di trú trong mùa đông khắc nghiệt, theo một cách không thể lường trước từ bắc Âu đến Baltic, Hà Lan, Anh và thậm chí tận nam Balkan.
Bạc má lớn đã được du nhập thành công vào Hoa Kỳ. Chúng được thả ở gần Cincinnati khoảng từ năm 1872 đến 1874 nhưng không thiết lập được tổ. Các kiến nghị cho rằng bạc má lớn là một biện pháp kiểm soát tuyệt vời đối với loài bướm đêm gây hại Cydia pomonella, dẫn đến việc du nhập chúng vào một số khu vực mới, đặc biệt là ở Hoa Kỳ. Tuy nhiên kế hoạch này đã không được thực hiện. Một quần thể nhỏ hiện diện ở thượng lưu Trung Tây được cho là hậu duệ của những cá thể chim được thả ở Chicago vào năm 2002 cùng với sẻ thông vàng châu Âu, quạ thông Á Âu, sẻ khướu, sẻ thông châu Âu, sẻ thông Saffron, sẻ ngô xanh và hồng tước, mặc dù một số loài có thể đã được du nhập trước. Bạc má lớn đã được nhập nội vào tỉnh Almaty (nay nằm ở Kazakhstan) vào những năm 1960–61 và đã thiết lập các tổ, dù tình trạng hiện tại của chúng không rõ lắm.
Hành vi
Chế độ ăn uống và cho ăn

Bạc má lớn chủ yếu ăn côn trùng và nhện mà chúng bắt được bằng cách mót tán lá vào mùa hè. Những con mồi không xương sống lớn hơn của loài bao gồm gián, châu chấu, dế, bọ cánh gân, bọ cánh da, bọ cánh nửa, kiến, ruồi, bọ cánh lông, bọ cánh cứng, ruồi bọ cạp, bọ chân dài, ong và ong bắp cày, ốc sên và bọ chân đều. Trong mùa sinh sản, bạc má lớn thường cho con ăn sâu bướm giàu protein. Một nghiên cứu được công bố vào năm 2007 cho thấy rằng bạc má lớn giúp giảm tới 50% thiệt hại do sâu bướm gây ra trong vườn táo. Chim non cũng được cho ăn một số loài nhện trong giai đoạn phát triển ban đầu, có thể vì lý do dinh dưỡng. Vào mùa thu và mùa đông, khi côn trùng trở nên khan hiếm hơn, bạc má lớn sẽ ăn thêm quả mọng và hạt. Hạt và quả thường là từ cây rụng lá và cây bụi, chẳng hạn như hạt của cây cử và cây trăn. Chim cũng sẽ lấy đồ ăn thừa trên bàn, hạt đậu phộng và hướng dương từ bàn chim, nếu có sẵn. Trong mùa đông cực kỳ khắc nghiệt, chúng ăn hạt hướng dương, loại thực phẩm chiếm 44% trọng lượng cơ thể. Chim thường kiếm ăn trên mặt đất, đặc biệt là vào những năm có số lượng hạt sồi cao. Bạc má lớn, cùng với những loài bạc má khác, sẽ tham gia vào đàn kiếm ăn hỗn hợp của các loài trong mùa đông.
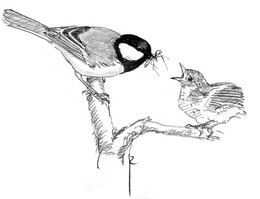
Các loại thực phẩm lớn, như hạt hoặc con mồi lớn, được đập vỡ, trong đó thức ăn được giữ bằng một hoặc cả hai chân và sau đó dùng mỏ đập nát cho đến khi có thể ăn được. Bằng cách này, bạc má lớn có thể đập vỡ một hạt phỉ trong khoảng 20 phút. Khi cho con non ăn, con trưởng thành sẽ loại bỏ đầu những con côn trùng lớn để dễ ăn hơn, đồng thời loại bỏ ruột côn trùng để tanin trong ruột không làm chim con chậm lớn.
Bạc má lớn kết hợp tính linh hoạt trong chế độ ăn uống với trí thông minh đáng kể và khả năng giải quyết vấn đề bằng cách học hỏi sâu sắc, tức là giải quyết vấn đề thông qua hiểu biết lớn hơn là thử và sai. Ở Anh, bạc má lớn đã học cách đập vỡ nắp giấy bạc của những chai sữa được giao trước cửa nhà để lấy phần kem trên cùng. Hành vi này được ghi nhận lần đầu tiên vào năm 1921, và đã lan rộng nhanh chóng trong hai thập kỷ tiếp theo. Vào năm 2009, người ta phát hiện những cá thể bạc má lớn đã giết và ăn não của những con dơi muỗi đang ngủ. Đây là lần đầu tiên một loài chim biết hót được ghi nhận có con mồi là dơi. Bạc má chỉ làm điều này trong mùa đông khi dơi đang ngủ đông và các loại thức ăn khác đang khan hiếm. Chúng cũng đã được ghi nhận là biết sử dụng các công cụ, như là dùng một cây kim bằng gỗ thông để lấy ấu trùng từ một cái lỗ trên cây.
Sinh sản
Bạc má lớn là loài sinh sản một vợ một chồng (bạn đời suốt đời) và thiết lập lãnh thổ sinh sản. Chim thiết lập các lãnh thổ này vào cuối tháng một, và bắt đầu xây dựng tổ vào cuối mùa đông hoặc đầu mùa xuân. Các lãnh thổ thường được tái sử dụng trong những năm liên tiếp, ngay cả khi một trong hai cá thể của cặp chết, miễn là đàn chim con được nuôi dưỡng thành công. Con cái có thể phân tán đến các lãnh thổ mới nếu tổ của chúng có trước năm trước. Nếu vì một lý do nào đó mà cặp đôi từ bỏ nhau thì đàn chim sẽ phân tán, với những con cái sẽ di chuyển xa hơn con đực để thiết lập lãnh thổ mới. Mặc dù bạc má lớn là loài một vợ một chồng về mặt xã hội, tình trạng lăng nhăng ở các cặp chim diễn ra thường xuyên. Một nghiên cứu ở Đức cho thấy 40% số tổ chứa một số con non do chim bố mẹ sinh sản chứ không phải do con đực khác, và 8,5% tổng số chim con là kết quả của việc cắm sừng. Những con đực trưởng thành có xu hướng sinh sản thành công hơn so với những con chưa trưởng thành.


Bạc má lớn là loài sinh sản theo mùa. Thời gian sinh sản chính xác thay đổi tùy theo một số yếu tố, quan trọng nhất là địa điểm. Mùa sinh sản hầu hết xảy ra từ tháng 1 đến tháng 9. Ở châu Âu, mùa sinh sản thường bắt đầu sau tháng 3. Ở Israel, có những ghi chép đặc biệt về việc sinh sản trong các tháng 10 đến tháng 12. Lượng ánh sáng mặt trời và nhiệt độ ban ngày cũng sẽ ảnh hưởng đến thời gian sinh sản. Một nghiên cứu đã tìm thấy mối tương quan chặt chẽ giữa thời điểm đẻ trứng và số lượng con mồi là sâu bướm, và điều này lại tương quan với nhiệt độ. Ở cấp độ cá thể, những con cái trẻ hơn có xu hướng bắt đầu đẻ muộn hơn những con cái lớn hơn.


Bạc má lớn là loài làm tổ trong hốc. Chúng thường sinh sản trong một cái lỗ bên trong cây, hoặc đôi khi là ở trên tường hoặc mặt đá, và chúng thường hay chui vào các hộp làm tổ. Con cái làm tổ bên trong hốc bằng sợi thực vật, cỏ, rêu, tóc, len và lông vũ. Số lượng trứng trong cùng một lần đẻ thường rất lớn, thường là từ 5 đến 12, và có thể lên đến 18. Số lượng trứng thấp hơn khi chim đẻ muộn hơn, hoặc khi mật độ đối thủ cạnh tranh cao hơn. Các cặp chim trẻ thường có số lượng trứng nhỏ hơn. Tính cách biệt cũng ảnh hưởng đến kích thước bầy, với những cá thể trên các đảo xa bờ đẻ trứng ít hơn và to hơn so với các các thể trên đất liền. Trứng có màu trắng với những đốm đỏ. Con cái đảm nhận việc ấp trứng và được con đực cho ăn trong quá trình ấp trứng. Bạc má lớn là loài chim gần gũi, rít lên khi bị quấy rầy. Thời điểm trứng nở được đồng bộ hóa tốt nhất với mức có sẵn con mồi cao nhất, có thể được điều chỉnh khi điều kiện môi trường thay đổi sau khi đẻ quả trứng đầu tiên bằng cách trì hoãn thời gian bắt đầu ấp, đẻ thêm trứng hoặc tạm dừng ấp. Thời gian ấp trứng từ 12 đến 15 ngày.

Giống như các loài bạc má khác, chim con nở ra không có lông và bị mù. Khi lông bắt đầu mọc ra, những cá thể con chậm phát triển hơn có bộ lông có màu carotenoid tương tự như bố mẹ của chúng (trong khi ở hầu hết các loài, bộ lông có màu xám để tránh bị ăn thịt). Gáy có màu vàng và có thể phản xạ tia cực tím để thu hút sự chú ý của chim bố mẹ. Điều này có thể giúp chúng dễ tìm thấy hơn trong điều kiện ánh sáng yếu, hoặc là một tín hiệu về thể trạng để thu hút sự chú ý của bố mẹ. Mảng màu sau gáy chuyển sang màu trắng sau lần thay lông đầu tiên khi được hai tháng tuổi, và thu hẹp lại khi chim lớn lên.
Chim con được cho ăn khoảng 6 đến 7 g (0,21–0,25 oz) thức ăn mỗi ngày. Cả chim bố và mẹ đều cho chim con ăn, và vệ sinh tổ bằng cách loại bỏ phân. Không có sự khác biệt về nỗ lực kiếm ăn giữa hai giới. Thời gian ở tổ của chim con kéo dài từ 16 đến 22 ngày, và độc lập với bố mẹ tám ngày sau khi biết bay. Việc cho chim non ăn có thể tiếp tục sau khi chim con độc lập, kéo dài đến 25 ngày ở lứa chim con đầu tiên, nhưng lên đến 50 ngày ở lứa thứ hai. Những cá thể con từ lứa thứ hai có hệ thống miễn dịch và tình trạng cơ thể yếu hơn so với những con từ lứa đầu tiên, do đó chim con có tỷ lệ sống sót thấp hơn.
Việc thoái hóa giống ở bạc má lớn xảy ra khi các con chim được sinh ra từ sự giao phối giữa các cá thể có quan hệ họ hàng gần. Sự suy giảm về thể lực thường được xem là kết quả của việc các alen có hại gia tăng biểu hiện ở các con chim này. Trong các cộng đồng tự nhiên của P. major, việc giao phối cận huyết được giảm bớt bằng cách phân tán các cá thể ra khỏi nơi sinh sống của chúng, giảm thiểu nguy cơ giao phối với họ hàng gần.
Sinh thái học
Cắt hỏa mai là một loài săn bạc má lớn. Những con con từ lứa thứ hai thường có nguy cơ bị tấn công cao hơn, một phần là do loài diều hâu này cần nhiều thức ăn hơn cho con non của chúng đang phát triển. Tổ của bạc má lớn thường bị kiến nhỏ sườn đỏ tấn công, đặc biệt là trong các loại tổ nhất định. Sóc xám miền Đông (nhập khẩu ở Anh) và triết bụng trắng cũng là những loài săn mồi của tổ, và chúng cũng có thể bắt các con trưởng thành làm tổ. Một loài rận (Mallophaga), được biết đến là Rostrinirmus hudeci, đã được phân lập và mô tả từ bạc má lớn ở Trung Âu vào năm 1981. Bọ chét gà mái Ceratophyllus gallinae rất phổ biến trong tổ của sẻ ngô xanh và bạc má lớn. Ban đầu chúng là một loài bọ chét chuyên dụng của bạc má, nhưng điều kiện khô ráo và đông đúc trong chuồng gà đã giúp chúng phát triển mạnh mẽ với vật chủ mới. Loài bọ chét này thường bị bọ hề Gnathoncus punctulatus săn. Ngoài ra, bọ cánh cứng Microglotta pulla cũng ăn bọ chét và ấu trùng của chúng. Mặc dù những con bọ cánh cứng này thường sống trong những tổ bỏ hoang, chúng chỉ có thể sinh sản ở nhiệt độ cao do sự ấp trứng của chim, và bạc má là loài vật chủ ưa thích. Những con bạc má lớn cạnh tranh với các con đớp ruồi châu Âu để giành được các hộp làm tổ, và có thể giết chết những con đớp ruồi đực. Các cuộc chiến tranh giành tồn vong diễn ra thường xuyên hơn khi thời gian làm tổ trùng khớp, và biến đổi khí hậu đã làm cho thời gian làm tổ của hai loài trở nên đồng bộ hơn, dẫn đến cái chết của đớp ruồi. Sau khi giết chết các con đớp ruồi, các con bạc má lớn có thể ăn não của chúng.
Sinh lý học
Bộ ngực lớn của loài chim được phát hiện có khả năng thích nghi sinh lý đặc biệt với môi trường lạnh. Chuẩn bị cho những tháng mùa đông, chim có thể tăng cường mức độ sinh nhiệt máu của chúng. Cơ chế thích ứng này bao gồm sự gia tăng theo mùa về thể tích ty thể và quá trình hô hấp của ty thể trong các tế bào hồng cầu, cùng với sự gia tăng tách rời chuỗi điện tử từ quá trình sản xuất ATP. Kết quả là năng lượng ban đầu được sử dụng để tạo ra ATP được giải phóng dưới dạng nhiệt và làm cho máu của chúng trở nên sinh nhiệt hơn. Trước tình trạng thiếu thốn thực phẩm vào mùa đông, bạc má lớn đã biểu hiện sự co mạch máu ngoại vi để giảm thiểu mất nhiệt và tổn thương do lạnh. Sự co mạch này được thực hiện thông qua sự lật ngược dòng máu của ngực lớn và sự co mạch ngoại vi ở các mạch chính xung quanh mỏ và chân của chim. Cơ chế này cho phép các vùng không được cách nhiệt (tức là mỏ và chân) vẫn duy trì nhiệt độ ổn định. Khi thiếu thực phẩm, nhiệt độ của mỏ chim giảm xuống, và khi thức ăn sẵn có tăng lên, nhiệt độ mỏ dần trở lại bình thường. Sự co mạch của các mạch máu trong mỏ không chỉ là một cơ chế tiết kiệm năng lượng, mà còn giúp giảm lượng nhiệt truyền từ các mô cơ thể chính đến da (thông qua sự giãn mạch ở da). Do đó, việc giảm tốc độ mất nhiệt bằng cách làm lạnh da so với môi trường làm giảm thiểu sự lãng phí năng lượng.
Mối quan hệ với con người

Bạc má lớn là một loài chim vườn phổ biến do khả năng nhào lộn của chúng khi ăn quả hạch hoặc hạt. Việc chim sẵn sàng di chuyển vào các hộp làm tổ đã khiến chúng trở thành một đối tượng nghiên cứu có giá trị trong ngành điểu học. Chúng đặc biệt hữu ích, vì giống như một mô hình để nghiên cứu sự tiến hóa của các đặc điểm lịch sử sự sống khác nhau, đặc biệt là kích thước ly hợp. Một nghiên cứu tìm kiếm tài liệu cơ sở dữ liệu đã tìm thấy 1.349 bài báo liên quan đến Parus major trong khoảng thời gian từ 1969 đến 2002.
Bạc má lớn nói chung đã thích nghi với những thay đổi của con người đối với môi trường. Chúng phổ biến hơn và sinh sản thành công hơn ở những khu vực có độ che phủ rừng không bị xáo trộn, nhưng cũng đã thích nghi với môi trường sống bị con người thay đổi. Chim có thể rất phổ biến ở các khu vực đô thị. Ví dụ, quần thể sinh sản ở thành phố Sheffield (một thành phố có nửa triệu dân) được ước tính vào khoảng 17.000 cá thể. Để thích nghi với môi trường có con người, tiếng hót của loài đã được quan sát thấy là thay đổi trong môi trường đô thị ô nhiễm tiếng ồn. Tiếng hót của loài có tần số cao hơn ở những khu vực có ô nhiễm tiếng ồn tần số thấp so với những khu vực yên tĩnh hơn. Loài bạc má này đã mở rộng phạm vi của chúng, di chuyển về phía bắc tới Scandinavia và Scotland, và phía nam tới Israel và Ai Cập. Tổng cá thể ước tính vào khoảng 300–1.100 triệu con chim trong phạm vi 32,4 triệu km (12,5 triệu dặm vuông Anh). Mặc dù đã có một số sự suy giảm dân số cục bộ ở những khu vực có môi trường sống kém chất lượng hơn, nhưng phạm vi rộng lớn và số lượng lớn của loài cho thấy bạc má lớn không bị coi là bị đe dọa, và chim được xếp là loài ít quan tâm trong Sách đỏ IUCN.