


| Bộ Đà điểu | |
|---|---|
| Thời điểm hóa thạch: Thế Paleocen-thế Holocen 56–0 triệu năm trước đây TiềnЄ
Є
O
S
D
C
P
T
J
K
Pg
N
| |
| Phân loại khoa học | |
| Giới (regnum) | Animalia |
| Ngành (phylum) | Chordata |
| Lớp (class) | Aves |
| Phân lớp (subclass) | Neornithes |
| Liên bộ (superordo) | Paleognathae |
| Bộ (ordo) | Struthioniformes Latham, 1790 |
| Các họ | |
Struthionidae (đà điểu châu Phi) | |
Bộ Đà điểu (Struthioniformes) bao gồm các loài chim lớn, không biết bay có nguồn gốc từ Gondwana, phần lớn trong số chúng đã tuyệt chủng. Khác với các loài chim không bay khác, các loài đà điểu không có xương chạc trên xương ức, dẫn đến việc thiếu điểm tựa chắc chắn cho cơ cánh, nên chúng không thể bay dù có cánh phù hợp để bay lượn.
Trước đây, các loài đà điểu chiếm ưu thế trên hầu hết các khu vực đại lục Gondwana, hoặc vẫn tồn tại cho đến thời gian gần đây.
Các loài hiện còn
- Đà điểu châu Phi là loài đà điểu lớn nhất còn tồn tại, có thể cao tới 3 m, nặng 135 kg và chạy nhanh hơn ngựa.
- Đà điểu Emu đứng thứ hai về kích thước, cao tới 2 m và nặng khoảng 60 kg. Tương tự như đà điểu châu Phi, nó chạy nhanh và là loài chim mạnh mẽ của các đồng bằng và khu rừng.
- Ba loài Đà điểu đầu mào có nguồn gốc từ Australia và các đảo phía bắc, ngắn hơn Emu và có cơ thể chắc chắn. Đà điểu đầu mào thích các khu rừng nhiệt đới rậm rạp và có thể rất nguy hiểm khi bị dồn vào thế bí hoặc bị bất ngờ. Ở New Guinea, trứng của đà điểu đầu mào được thu hoạch và nở thành con non để ăn thịt, dù có thể gây rủi ro cho con người.
- Ở New Zealand, có 5 loài chim kiwi nhỏ nhất, kích thước tương đương gà, nhút nhát và dễ cáu giận. Chúng sống trong các hang sâu và dùng khứu giác nhạy bén để tìm kiếm côn trùng nhỏ trong đất. Kiwi đặc biệt vì trứng của chúng chiếm tỷ lệ rất lớn so với trọng lượng cơ thể của chim mái, có thể đạt 15-20% trọng lượng cơ thể.
- Nam Mỹ có 2 loài đà điểu Nam Mỹ có kích thước trung bình, nhanh nhẹn trên các cánh đồng cỏ (pampa). Các cá thể lớn nhất của loài đà điểu Nam Mỹ lớn (Rhea americana) có thể cao tới 1,5 m và nặng 20 – 25 kg. Ngoài ra, Nam Mỹ còn có 47 loài thuộc 9 chi trong họ Tinamidae, bao gồm các loài chim nhỏ, sống dưới đất nhưng không phải không bay được, và có họ hàng gần với các loài đà điểu.
Những loài đã bị tuyệt chủng
- Aepyornis, hay còn gọi là 'chim voi' ở Madagascar, từng là loài chim lớn nhất được biết đến. Mặc dù chúng thấp hơn so với các loài moa cao nhất, nhưng những cá thể lớn nhất có thể nặng tới 450 kg. Hai loài đã tồn tại khi con người di cư đến từ Borneo và châu Phi, có thể từ thế kỷ I. Cả hai dường như sống sót khá lâu: loài Aepyornis mullerornis nhỏ hơn có thể đã tuyệt chủng sớm hơn, trong khi loài Aepyornis maximus lớn hơn có thể còn tồn tại đến đầu thế kỷ XVII.
- Họ Dinornithidae (moa) có ít nhất 11 loài khác nhau từng sống tại New Zealand cho đến khi con người đến đây vào thế kỷ XIII hoặc sớm hơn. Chúng có kích thước từ như gà tây đến moa khổng lồ (Dinornis giganteus) với chiều cao lên đến 3,3 m (11 ft) và trọng lượng tới 250 kg (550 lb). Giống như đà điểu đầu mào, moa sống chủ yếu trong các khu rừng mà không có kẻ thù là động vật ăn thịt. Chúng được cho là đã tuyệt chủng khoảng năm 1500 do sự săn bắn kéo dài vài trăm năm kể từ khi có sự định cư của con người. Tuy nhiên, một số người vẫn tin rằng có thể còn tồn tại những quần thể nhỏ trong các khu vực hoang vắng cho đến gần đây.
Ngoài ra, các mảnh vỏ trứng giống như vỏ trứng của Aepyornis cũng đã được phát hiện trên quần đảo Canary. Các mảnh này có niên đại từ Trung hoặc Hậu Miocen, và không có giả thuyết hợp lý nào về cách chúng đến đây, do không chắc chắn về việc các đảo này có từng nối liền với đại lục hay không.
Tiến hóa và phân loại
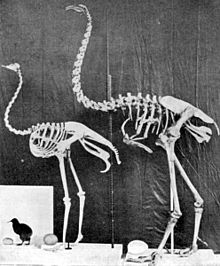
Hiện nay có hai phương pháp phân loại chim chạy: phương pháp đầu tiên, được áp dụng trong bài này, nhóm các họ thuộc bộ Struthioniformes thành một nhóm; trong khi phương pháp thứ hai cho rằng các dòng dõi đã tiến hóa độc lập và do đó nâng các họ lên thành các bộ riêng biệt (như Rheiformes, Casuariiformes, Apterygiformes, v.v.).
Một số nghiên cứu dựa trên hình thái học, miễn dịch học và trình tự DNA cho thấy các loài chim chạy là đơn ngành. Theo mô hình truyền thống, chim chạy là nhóm chim không bay xuất hiện ở Gondwana trong kỷ Creta, sau đó phát triển tách biệt khi các lục địa phân tách. Tuy nhiên, phân tích gần đây về biến thiên gen giữa các loài chim chạy cho thấy chúng tách ra từ nhau gần đây hơn so với tổ tiên Gondwana chung. Thêm vào đó, hóa thạch Trung Eocen của 'tiền-đà điểu' Palaeotis từ Trung Âu có thể gợi ý rằng giả thuyết 'ngoài Gondwana' là không chính xác. Phân tích gần đây với 20 gen hạt nhân cũng đặt câu hỏi về tính đơn ngành của nhóm, gợi ý rằng các loài tinamou có thể cũng thuộc vào dòng dõi chim chạy.
Nghiên cứu so sánh toàn bộ trình tự DNA ti thể của các loài chim chạy còn tồn tại và 2 loài moa (khủng điểu) chỉ ra rằng moa là nhóm cơ sở, tiếp theo là đà điểu châu Mỹ, sau đó là đà điểu châu Phi, kiwi, và cuối cùng là đà điểu Úc (emu) và đà điểu đầu mào là các họ hàng gần nhất. Một nghiên cứu khác đảo ngược vị trí tương đối của moa và đà điểu châu Mỹ, chỉ ra rằng chim voi không phải là họ hàng gần của đà điểu châu Phi hay các loài chim chạy khác. Nghiên cứu gen hạt nhân cho thấy đà điểu châu Phi tách ra đầu tiên, tiếp theo là đà điểu châu Mỹ và tinamou, sau đó là kiwi phân tách từ đà điểu Úc và đà điểu đầu mào. Các nghiên cứu cho thấy tổ tiên của moa có thể đã tồn tại ở New Zealand từ khi tách khỏi Gondwana, trong khi tổ tiên của kiwi có thể đã di cư từ Australia gần đây hơn, có thể qua cầu đất liền hoặc bằng cách 'nhảy' qua các đảo. Theo phân tích trước đây, đà điểu châu Phi có thể đã tới châu Phi sau khi tách khỏi Nam Mỹ (như qua xâm lấn đại lục Á Âu và sau đó châu Phi tách khỏi Ấn Độ), nhưng dữ liệu hạt nhân cho thấy đà điểu châu Phi có lẽ tách ra đầu tiên, phù hợp với trình tự tách mảng kiến tạo của Gondwana. Nhiều khía cạnh trong cổ địa sinh học của chim chạy phù hợp với giả thuyết hình thành loài theo địa lý (sự chia tách kiến tạo mảng của Gondwana).
Các nghiên cứu gần đây về bộ gen phát sinh loài cho thấy tinamou có thể thực sự thuộc về nhóm chim chạy. Nếu đúng như vậy, nhóm 'chim chạy' trở nên cận ngành thay vì đơn ngành. Điều này đặt ra câu hỏi đáng chú ý về tiến hóa khả năng bay và không bay trong nhóm này, bởi vì chim chạy truyền thống được cho là có tổ tiên không bay và là nhóm đơn ngành, trong khi sự phân tách của tinamou trong dòng dõi chim chạy gợi ý rằng khả năng bay có thể đã tái phát triển ở tinamou hoặc đã mất đi ở các loài chim chạy khác.
| Biểu đồ phát sinh chủng loài vẽ theo Mitchell et al. (2014) | |||||||||||||||||||||||||||||||||||||||||||||
|
Vào năm 2014, một phân tích DNA ti thể bao gồm cả các thành viên hóa thạch cho thấy tinamou nằm sâu trong nhóm đà điểu. Đà điểu châu Phi được xếp ở nhánh cơ sở, tiếp theo là đà điểu Nam Mỹ, rồi đến nhánh chứa moa và tinamou, sau đó là hai nhánh cuối cùng: một nhánh chứa emu và đà điểu đầu mào, nhánh còn lại chứa chim voi và kiwi. Mối quan hệ giữa moa và tinamou phù hợp với các phát hiện trước đây, trong khi mối quan hệ giữa chim voi và kiwi là phát hiện mới. Phân tích hình thái học cũng hỗ trợ thêm cho kết quả này.
Ảnh hưởng đến nền văn minh nhân loại
Trong thời kỳ Ai Cập cổ đại, lông đà điểu được sử dụng để biểu thị ý nghĩa chính nghĩa vì các sợi lông của nó khá đều nhau.
Các loài chim trong tưởng tượng
Trong loạt trò chơi điện tử Final Fantasy, Chocobo là loài chim tưởng tượng tương tự như chim chạy. Trong các trò chơi này, chocobo được dùng như phương tiện di chuyển, có thể cưỡi hoặc kéo xe như ngựa.
Một loài chim đáng chú ý là roc (từ tiếng Ba Tư: رخ - rokh), được miêu tả trong truyền thuyết Ả Rập trung cổ như một loài chim khổng lồ có thể nâng cả voi. Theo một số nguồn, roc có thể chính là đà điểu châu Phi. Nguồn gốc của quân xe trong cờ vua có thể bắt nguồn từ đây.
Hình ảnh


Ghi chú và Các liên kết bên ngoài
Chim (lớp: Aves) |
|---|
| Tiêu đề chuẩn |
|
|---|
| Thẻ nhận dạng đơn vị phân loại |
|---|