
Trong di truyền học, bộ gen (phát âm IPA: /əʊ piːrɒn/, tiếng Việt: ô-pê-ron) là một nhóm gen cấu trúc liền kề trên DNA của sinh vật nhân sơ và virus, có cùng một cơ chế điều hòa.
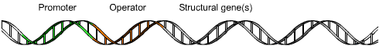
Về mặt chức năng, mỗi bộ gen là một đơn vị hoạt động của phân tử DNA, bao gồm một hoặc nhiều gen cấu trúc (cistron) liền kề nhau, có cùng một vùng điều hòa (operator) và một vùng khởi động (promoter) duy nhất.
Đặc điểm
- Khi hoạt động (bộ gen mở) thì các gen trong bộ gen cùng được phiên mã với nhau thành một chuỗi phân tử mRNA ghép, gọi là RNA đa xi-trôn (polycistronic mRNA). Sau khi được phiên mã, chúng có thể được dịch mã cùng nhau trong bào tương, hoặc trải qua chế biến để tạo ra các mRNA đơn xi-trôn rồi được dịch mã riêng biệt. Do đó, kết quả là các gen cùng một bộ gen thường được biểu hiện cùng lúc hoặc không được biểu hiện.
- Khi bộ gen đóng, thì sản phẩm (là mRNA và sau đó là protein) không được tạo ra, do đó bộ gen là một đơn vị dùng để điều hoà hoạt động gen chỉ ở cấp độ phiên mã.
Lịch sử
Thuật ngữ 'bộ gen' được đề xuất và sử dụng đầu tiên bởi hai nhà di truyền học người Pháp là François Jacob và Jacques Monod trong các nghiên cứu của họ về vi khuẩn E. coli. Có tác giả cho rằng thuật ngữ này xuất hiện lần đầu tiên trong một bài báo ngắn ở Kỷ yếu Viện Hàn lâm Khoa học Pháp vào năm 1960. Từ bài báo này, lý thuyết chung của bộ gen có thể nói là đã được phát triển. Lý thuyết này gợi ý rằng trong mọi trường hợp, các gen trong một bộ gen được kiểm soát âm tính bởi một chất ức chế hoạt động ở một operator (vùng vận hành) duy nhất nằm trước gen đầu tiên. Sau đó, người ta phát hiện ra rằng các gen cũng có thể được điều hòa dương tính cũng như có thể được điều chỉnh ở các bước sau, chứ không chỉ ở bước khởi đầu phiên mã. Do đó, không thể nói rằng tất cả bộ gen có một cơ chế điều hòa chung, vì các bộ gen khác nhau lại có các cơ chế khác nhau. Ngày nay, bộ gen được định nghĩa đơn giản là: một nhóm gen được phiên mã thành một phân tử mRNA duy nhất. Tuy nhiên, việc phát triển của lý thuyết bộ gen được coi là một sự kiện mang tính bước ngoặt trong lịch sử ngành sinh học phân tử. Bộ gen đầu tiên được miêu tả là operon lac ở vi khuẩn E. coli. Giải Nobel Sinh lý và Y học năm 1965 được trao cho François Jacob, André Michel Lwoff và Jacques Monod vì những phát hiện của họ liên quan đến sự tổng hợp operon và virus.
Ban đầu, các bộ gen được cho là chỉ tồn tại trong sinh vật nhân sơ (bao gồm các bào quan như lạp thể có nguồn gốc từ vi khuẩn), nhưng kể từ khi phát hiện ra những bộ gen đầu tiên trong sinh vật nhân chuẩn vào đầu những năm 1990, chúng phổ biến hơn so với giả định trước đây. Nói chung, sự biểu hiện của các bộ gen tạo ra các mRNA đa cistron, trong khi các bộ gen nhân chuẩn nhân tạo ra các mRNA đơn cistron.
Bộ gen cũng được tìm thấy trong các loại virus như virus thực khuẩn. Ví dụ, các phage T7 có hai bộ gen. Bộ gen đầu tiên cho các sản phẩm khác nhau, bao gồm cả RNA polymerase T7 đặc biệt có thể liên kết và phiên mã cho bộ gen thứ hai. Bộ gen thứ hai bao gồm một gen làm tan có nghĩa là khiến cho tế bào bị phá hủy. Ở nhóm này, hệ thống điều hòa di truyền được tìm thấy ở vi khuẩn và virus của chúng trong đó các gen mã hóa cho các protein liên quan đến chức năng được tập hợp dọc theo DNA. Tính năng này cho phép tổng hợp protein được kiểm soát phối hợp để đáp ứng nhu cầu của tế bào. Bằng cách cung cấp các phương tiện để sản xuất protein chỉ khi và khi chúng được yêu cầu, bộ gen cho phép tế bào bảo tồn năng lượng (là một phần quan trọng trong chiến lược cuộc sống của sinh vật). Một bộ gen điển hình bao gồm một nhóm các gen cấu trúc mã hóa các enzyme tham gia vào quá trình trao đổi chất, chẳng hạn như sinh tổng hợp một amino acid. Những gen này nằm liền kề nhau trên một đoạn DNA và nằm dưới sự kiểm soát của một nhà tổ chức (một đoạn DNA ngắn mà RNA polymerase liên kết để bắt đầu phiên mã). Một đơn vị RNA thông tin (mRNA) được phiên mã từ bộ gen và sau đó được dịch thành các protein riêng biệt.
Ôpêron phổ biến chủ yếu trong sinh vật nhân sơ nhưng cũng có mặt ở một số sinh vật nhân chuẩn như giun tròn như C. elegans và ruồi giấm, Drosophila melanogaster. Gen rRNA thường tồn tại trong các ôpêron đã được tìm thấy ở nhiều sinh vật nhân chuẩn bao gồm cả động vật có xương sống. Một ôpêron được tạo thành từ một số gen cấu trúc được sắp xếp dưới sự điều khiển của một promoter chung và được điều hòa bởi một operator (vùng vận hành) chung. Ôpêron được định nghĩa là một tập hợp các gen cấu trúc liền kề, kèm theo các tín hiệu điều hòa lân cận ảnh hưởng đến sự phiên mã của các gen cấu trúc. Các yếu tố điều hòa của một ôpêron nhất định, bao gồm cả chất ức chế, chất đồng ức chế và chất hoạt hóa, không nhất thiết phải mã hóa bởi ôpêron. Vị trí và điều kiện của các chất điều hòa, promoter, operator và các chuỗi DNA cấu trúc có thể xác định tác động của các đột biến phổ biến.
Các ôpêron liên quan đến các thuật ngữ như regulon và stimulon; trong khi ôpêron chứa một tập hợp các gen được điều khiển bởi cùng một operator, các regulon chứa một tập hợp các gen được điều hòa bởi một protein điều hòa duy nhất (regulate), và các stimulon chứa một tập hợp các gen được điều hòa bởi một kích thích tế bào đơn (stimulus). Theo các tác giả, thuật ngữ 'ôpêron' có nguồn gốc từ động từ 'operate' (vận hành).
Tất cả các gen cấu trúc của một ôpêron đều được BẬT hoặc TẮT cùng nhau, điều này có được nhờ vào một promoter và operator nằm ngược dòng với chúng, nhưng đôi khi cần nhiều hơn thế để có thể kiểm soát biểu hiện gen. Để đạt được khía cạnh này, một số gen vi khuẩn được đặt gần nhau, nhưng có một promoter cụ thể cho từng gen trong số này; điều này được gọi là phân nhóm gen. Thông thường những gen này mã hóa cho các protein sẽ hoạt động cùng nhau trong cùng một con đường, chẳng hạn như là con đường trao đổi chất. Phân nhóm gen giúp tế bào nhân sơ có thể tạo ra các enzyme chuyển hóa theo đúng thứ tự.
Cấu trúc tổng quát
Một ôpêron bao gồm ba thành phần cơ bản:
- Promoter là vùng khởi động - thực chất là một chuỗi nucleotide nhất định để enzyme RNA polymerase nhận diện và gắn vào. RNA polymerase có đơn vị nhỏ chứa trình tự nucleotide tương ứng với vùng khởi động này. Dù operon mở hay đóng, enzyme vẫn nhận diện và gắn vào vùng này, nhưng chỉ hoạt động khi operon mở, sau đó bắt đầu phiên mã. Trong quá trình phiên mã tạo ra RNA, vùng khởi động (promoter) chỉ ra gen nào sẽ được sử dụng để tạo ra mRNA (RNA tin tức), từ đó điều khiển sản xuất protein trong tế bào.
- Operator là vùng vận hành - thực chất là một chuỗi nucleotide nhất định trên DNA nằm ngay sau vùng khởi động. Nếu có một chất (thường là protein) liên kết vào vùng này, operon sẽ bị đóng, vì vậy chất này được gọi là chất ức chế (repressor). Nguyên nhân đơn giản của việc 'đóng cửa' operon là chất ức chế đã chặn đường (giống như một chiếc barie), ngăn không cho RNA polymerase trượt dọc theo nhóm gen trong operon. Nói một cách khác, chất ức chế làm gián đoạn hoạt động vật lí của phân tử RNA polymerase.
- Các gen cấu trúc - các gen được điều hòa bởi operon.
Không phải lúc nào cũng bao gồm gen điều hòa trong operon, nhưng một gen điều hòa là quan trọng trong chức năng của operon, nó là một gen được biểu hiện liên tục mã hóa cho một protein ức chế. Gen điều hòa không cần phải liền kề hoặc gần operon để kiểm soát nó.
Hoạt động
Kiểm soát operon là một loại điều hòa gen cho phép sinh vật điều chỉnh sự biểu hiện của các gen khác nhau tùy thuộc vào điều kiện môi trường. Cơ chế hoạt động có thể là cảm ứng hoặc ức chế, tích cực hoặc tiêu cực tương ứng với mỗi loại này.
Kiểm soát âm tính liên quan đến sự kiện một chất ức chế liên kết vào operator để ngăn cản phiên mã.
- Trong các operon cảm ứng âm tính, một protein ức chế mặc định liên kết với operator, ngăn cản phiên mã của các gen trong operon. Khi có phân tử cảm ứng xuất hiện, nó sẽ kết hợp với protein ức chế này và thay đổi cấu trúc của nó để không thể liên kết với operator nữa. Điều này cho phép operon bắt đầu biểu hiện. Operon lac là một ví dụ về operon cảm ứng âm tính, trong đó phân tử cảm ứng là allolactose.
- Ở các operon ức chế âm tính, phiên mã của operon diễn ra theo mặc định. Protein ức chế được sản sinh từ gen điều hòa, nhưng chúng không thể liên kết với operator trong trạng thái cấu hình bình thường. Tuy nhiên, một số phân tử đặc biệt gọi là chất đồng ức chế sẽ kết hợp với protein ức chế, thay đổi cấu trúc và kích hoạt hoạt tính ức chế của chúng. Protein ức chế hoạt động sẽ liên kết với operator và ngăn cản phiên mã. Operon trp, tham gia vào quá trình tổng hợp tryptophan (và chính allolactose đóng vai trò là chất đồng ức chế), là một ví dụ về operon điều hòa theo cơ chế ức chế âm tính.
Các operon cũng có thể được điều hòa dương tính. Trong điều hòa dương tính, một protein hoạt hóa kích thích phiên mã bằng cách liên kết với DNA (thường là tại một vị trí khác với operator).
- Trong các operon cảm ứng dương tính, các protein hoạt hóa thường không thể liên kết với DNA. Khi một phân tử cảm ứng kết nối với protein hoạt hóa, protein sẽ thay đổi cấu trúc để có thể kết nối với DNA và kích hoạt phiên mã.
- Ở các operon ức chế dương tính, các protein hoạt hóa mặc định có thể liên kết với một đoạn DNA phù hợp. Tuy nhiên, khi một chất ức chế kết nối với protein hoạt hóa này, protein sẽ không thể liên kết với DNA nữa. Điều này ngăn chặn hoạt hóa và ngăn cản việc phiên mã trong hệ thống.
Operon lac
Operon lac của vi khuẩn mô hình Escherichia coli là operon đầu tiên được phát hiện và cung cấp cho chúng ta một ví dụ điển hình về chức năng của operon. Operon này gồm ba gen cấu trúc nằm kế nhau, một bộ điều hòa, một điểm kết thúc và một toán tử. Các operon lac được điều chỉnh bởi nhiều yếu tố, trong đó có sẵn glucose và không có sẵn lactose. Operon có thể được kích hoạt bởi allolactose. Lactose gắn vào protein ức chế và làm giảm ức chế sự phát mã của gen. Đây là một ví dụ về mô hình ức chế gen (nếu phân loại như trên: cảm ứng âm tính).
Operon trp
Operon trp được phát hiện vào năm 1953 bởi Jacques Monod và đồng nghiệp, là operon ức chế đầu tiên được phát hiện trong E. coli. Trong khi operon lac có thể được kích hoạt bởi một chất hóa học (allolactose), thì operon tryptophan (Trp) lại bị ức chế bởi một chất hóa học (tryptophan). Operon này bao gồm năm gen cấu trúc: trp E, trp D, trp C, trp B và trp A, mã hóa cho enzyme tryptophan synthetase. Nó cũng bao gồm một bộ điều hòa kết nối với RNA polymerase và một toán tử ngăn chặn sự phát mã, nếu protein được tổng hợp bởi gen ức chế (trp R) liên kết với vùng toán tử này. Nếu trong operon lac, lactose gắn vào protein ức chế và giảm ức chế phát mã gen, thì trong operon trp, tryp...
Dự đoán số lượng và tổ chức của các operon
Số lượng và tổ chức của các operon đã được nghiên cứu kỹ lưỡng nhất trong E. coli. Kết quả là, chúng ta có thể dự đoán dựa trên trình tự gen của sinh vật.
Một phương pháp dự đoán sử dụng khoảng cách giữa các khung đọc như một yếu tố quan trọng trong việc dự đoán số lượng operon trong hệ gen. Quá trình tách biệt chỉ là thay đổi khung đọc và đảm bảo rằng quá trình 'đọc' là hiệu quả. Những khoảng cách giữa các operon thường dao động từ 40 đến 50 base.
Một phương pháp thay thế để dự đoán operon là tìm kiếm các cụm gen nơi thứ tự và định hướng gen được bảo tồn trong hai hoặc nhiều bộ gen.
Dự đoán operon có thể chính xác hơn nếu xem xét các chức năng của các phân tử. Vi khuẩn tổ chức khung đọc của chúng thành các đơn vị, với sự tham gia của các phức hợp protein, con đường chung hoặc các cơ chất và protein vận chuyển. Do đó, dự đoán chính xác sẽ phụ thuộc vào tất cả các dữ liệu này, thật sự là một nhiệm vụ khó khăn.
Phòng thí nghiệm của Pascale Cossart là phòng thí nghiệm đầu tiên xác định tất cả các operon của vi khuẩn Listeria monocytogenes. Nghiên cứu năm 2009 mô tả 517 operon đa cistron và các thay đổi tổng hợp trong phiên mã của L. monocytogenes dưới các điều kiện khác nhau.