

| Dilophosaurus | |
|---|---|
| Khoảng thời gian tồn tại: Jura sớm, (tầng Sinemur), 193 triệu năm trước đây TiềnЄ
Є
O
S
D
C
P
T
J
K
Pg
N
↓ | |
| Mô hình mẫu định danh (UCMP 37302) trong tư thế nguyên trạng, Bảo tàng Royal Ontario | |
| Phân loại khoa học | |
| Vực: | Eukaryota |
| Giới: | Animalia |
| Ngành: | Chordata |
| Lớp: | Reptilia |
| nhánh: | Dinosauria |
| Bộ: | Saurischia |
| Phân bộ: | Theropoda |
| Họ: | †Dilophosauridae |
| Chi: | †Dilophosaurus Welles, 1970 |
| Loài: | †D. wetherilli
|
| Danh pháp hai phần | |
| †Dilophosaurus wetherilli (Welles, 1954) | |
| Các đồng nghĩa | |
| |
Dilophosaurus (phát âm tiếng Anh: /daɪˌloʊfəˈsɔːrəs,
Với chiều dài khoảng 7 mét và trọng lượng khoảng 400 kg, Dilophosaurus là một trong những loài khủng long ăn thịt sớm nhất, mặc dù nó nhỏ hơn nhiều so với các loài khủng long chân thú sau này. Nó có cơ thể mảnh mai và nhẹ, với hộp sọ lớn nhưng tinh tế. Mõm của nó hẹp, và hàm trên có một khoảng trống hoặc kẽ hở bên dưới lỗ mũi. Nó có một cặp mào hình đĩa trên đầu, giống như đà điểu đầu mào nhưng có hai đỉnh. Xương hàm dưới mảnh mai ở phía trước nhưng sâu ở phía sau. Răng dài, cong, mỏng và bị nén ngang. Răng ở hàm dưới nhỏ hơn nhiều so với hàm trên. Hầu hết các răng đều có răng cưa ở hai mặt. Cổ dài với các đốt sống rỗng và nhẹ. Các chi cánh tay mạnh mẽ, với xương cánh tay dài và mảnh mai. Bàn tay có bốn ngón tay: ngón đầu ngắn nhưng mạnh mẽ với móng vuốt lớn, hai ngón tay sau dài hơn và mảnh hơn với móng vuốt nhỏ hơn, và ngón thứ tư là di tích. Xương đùi lớn và bàn chân mạnh mẽ với móng vuốt lớn.
Dilophosaurus là thành viên của họ Dilophosauridae, cùng với Dracovenator, một nhóm nằm giữa Coelophysidae và các theropod sau này. Dilophosaurus là loài khủng long đi bằng hai chân, có thể đã săn các loài động vật lớn, nhưng cũng ăn các loài nhỏ hơn và cá. Chức năng của các mào trên đầu chưa được xác định rõ; chúng có thể không đủ mạnh để chiến đấu, nhưng có thể được sử dụng để phô trương, nhận diện loài, hoặc lựa chọn bạn tình. Nó có thể phát triển nhanh chóng, đạt tốc độ tăng trưởng từ 30 đến 35 kg mỗi năm vào giai đoạn đầu đời. Mẫu vật gốc có nhiều dấu hiệu bệnh lý, bao gồm các chấn thương đã lành và dị thường phát triển. Dilophosaurus được biết đến từ Thành hệ Kayenta và sống chung với các loài khủng long như Megapnosaurus và Sarahsaurus. Dilophosaurus đã được xuất hiện trong tiểu thuyết Jurassic Park và phim chuyển thể cùng tên, nơi chúng được miêu tả với khả năng hư cấu như phun nọc độc và xòe hai cái bờm, cùng với kích thước nhỏ hơn thực tế. Loài này được chỉ định làm loài khủng long biểu trưng của bang Connecticut vào năm 2017.
Lịch sử nghiên cứu
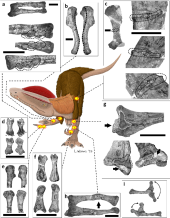
Vào mùa hè năm 1942, nhà cổ sinh vật học Charles L. Camp từ Đại học Bảo tàng cổ sinh vật học California (UCMP) dẫn một nhóm nghiên cứu đi tìm kiếm hóa thạch ở miền Bắc quận Navajo, Arizona. Tin tức về chuyến khảo sát đã thu hút sự chú ý của cộng đồng người Mỹ bản địa, và Jesse Williams, một người Navajo, đã dẫn ba thành viên của đoàn đến nơi ông phát hiện vài mảnh xương hóa thạch vào năm 1940. Khu vực này thuộc thành hệ Kayenta, cách khoảng 32 km về phía bắc Cameron, gần thành phố Tuba trong Xứ Navajo. Ba bộ xương khủng long đã được tìm thấy trong đá phiến sét màu tía, tạo thành một hình tam giác với cạnh dài khoảng 9,1 m. Bộ xương đầu tiên gần như hoàn chỉnh, chỉ thiếu phần đầu sọ, xương chậu và một số đốt sống. Bộ xương thứ hai bị hư hại nghiêm trọng, chỉ còn mặt trước của hộp sọ, hàm dưới, một số đốt sống, xương chân tay, và một bàn tay vẫn còn nguyên. Bộ xương thứ ba bị bào mòn nhiều, chỉ còn vài đốt xương sống. Bộ xương hoàn chỉnh nhất được bao bọc trong thạch cao sau 10 ngày khai quật vất vả và được chất lên xe tải, trong khi bộ xương thứ hai được thu thập dễ dàng vì lộ thiên gần như hoàn toàn, còn bộ xương thứ ba gần như mất tích.
Quá trình làm sạch và lắp ráp bộ xương thứ nhất được thực hiện bởi nhóm ba nhà khoa học tại UCMP dưới sự giám sát của nhà cổ sinh vật học Wann Langston Jr., và mất tới hai năm để hoàn tất. Bộ xương được lắp trên tường với một số thay đổi nhỏ: đuôi được cong lên trên, cổ được duỗi thẳng hơn, và chân trái được nâng lên để dễ nhìn hơn. Do hộp sọ mẫu gốc bị nát, các nhà khoa học phải tái tạo nó bằng cách kết hợp mặt sau của mẫu đầu tiên và mặt trước của mẫu thứ hai. Xương chậu được tái tạo dựa trên mẫu Allosaurus, và các bàn chân cũng cần phục dựng lại. Đây là bộ xương được bảo tồn tốt nhất trong số các khủng long chân thú thời bấy giờ, dù vẫn chưa hoàn chỉnh. Năm 1954, Samuel P. Welles, một thành viên của nhóm khai quật, mô tả sơ bộ và định danh loài này là một loài mới thuộc chi Megalosaurus, đặt tên là M. wetherilli. Mẫu vật gần hoàn chỉnh (UCMP 37302) trở thành mẫu định danh, còn mẫu thứ hai (UCMP 37303) là mẫu phụ. Danh pháp loài được đặt theo tên của John Wetherill, một ủy viên hội đồng Navajo, mà Welles miêu tả là 'nhà thám hiểm, bạn của các nhà khoa học, và nhà giao dịch đáng tin cậy'. Cháu trai của Wetherill, Milton, là người đã thông báo về hóa thạch cho đoàn khảo sát. Welles xếp loài này vào chi Megalosaurus vì tỷ lệ cơ thể tương tự với M. bucklandii và không có sự khác biệt rõ rệt nào khác. Vào thời điểm đó, Megalosaurus được coi là 'thùng rác phân loại', nơi nhiều loài khủng long bị phân loại sơ sài được đưa vào.
Welles trở lại thành phố Tuba vào năm 1964 để xác định niên đại của thành hệ Kayenta (mặc dù có ý kiến cho rằng nó thuộc kỷ Trias muộn, nhưng Welles cho rằng nó thuộc kỷ Jura sớm hoặc giữa), và phát hiện một bộ xương khác cách 402,3 km về phía nam so với các mẫu vật tìm được năm 1942. Mẫu xương mới này gần như hoàn chỉnh (UCMP 77270), được thu thập với sự hỗ trợ của William Breed từ Bảo tàng Bắc Arizona và nhiều cộng sự. Trong quá trình chuẩn bị, họ nhận thấy mẫu này lớn hơn hẳn so với các mẫu M. wetherilli trước đó, và có hai cái mào trên đầu. Do các mào khá mỏng, ban đầu các nhà nghiên cứu tưởng nhầm chúng là mảnh xương của mái sọ bị loài ăn xác làm mất. Sau khi nhận ra đó là các mào nằm ở bên phải đường giữa, họ cũng phát hiện một cái mào đối xứng bên trái. Phát hiện này khiến họ phải kiểm tra lại mẫu định danh và tìm thấy phần cuống còn sót lại của hai mảnh xương mỏng hướng lên trên, bị nghiền nát với nhau. Trước đây, chúng được coi là hai phần của xương gò má bị thất lạc. Họ kết luận rằng các mẫu vật năm 1942 đều là các con non, trong khi mẫu năm 1964 là con trưởng thành, lớn hơn khoảng một phần ba so với các mẫu trước.


Welles và một trợ lý đã chỉnh sửa giá treo của mẫu gốc dựa trên bộ xương mới, phục dựng lại mào và xương chậu, kéo dài xương sườn cổ, và đặt chúng gần nhau hơn. Sau khi nghiên cứu các bộ xương khủng long chân thú ở Bắc Mỹ và châu Âu, Welles nhận ra rằng loài này không thuộc về chi Megalosaurus, và cần phải tạo một cái tên chi mới. Vào thời điểm đó, chưa có mẫu loài khủng long nào với cặp mào trên đầu như vậy, do đó giới cổ sinh vật học rất quan tâm. Một bản sao của mẫu gốc được tạo dựng và các phiên bản sợi thủy tinh đã được phân phối cho các triển lãm; để dán nhãn cho các mô hình, Welles quyết định đặt tên chi mới trong một ghi chú ngắn gọn thay vì chờ công bố chi tiết. Năm 1970, Welles đặt tên chi mới là Dilophosaurus, từ tiếng Hy Lạp di (δι) nghĩa là 'hai', lophos (λόφος) nghĩa là 'đỉnh' hay 'mào', và sauros (σαυρος) nghĩa là 'thằn lằn'; ghép lại thành 'thằn lằn hai mào'. Welles công bố mô tả chi tiết về bộ xương Dilophosaurus vào năm 1984, nhưng mẫu năm 1964 vẫn chưa được mô tả chính thức. Dilophosaurus là loài khủng long chân thú được nghiên cứu kỹ nhất từ tầng đá Jura sớm và cũng thường là những bộ xương được bảo tồn tốt nhất ở độ tuổi đó.
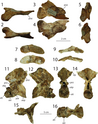
Năm 2001, nhà cổ sinh vật học Robert J. Gay xác định ít nhất ba mẫu vật Dilophosaurus mới (dựa trên sự hiện diện của ba mảnh xương mu và hai khuỷu tay khác nhau) trong bộ sưu tập của Bảo tàng Bắc Arizona. Các mẫu vật này được tìm thấy vào năm 1978 tại Rock Head Quadrangle, cách 190 km so với nơi các mẫu vật gốc được phát hiện, và đều được dán nhãn là 'khủng long chân thú lớn'. Mặc dù phần lớn các vật liệu bị hư hại, chúng vẫn là bổ sung quan trọng cho mẫu gốc, bao gồm một phần xương chậu và một số xương sườn. Một số mẫu thuộc về một con non (MNA P1.3181), là mẫu vật trẻ nhất của chi này và là một trong những khủng long non được khai quật sớm nhất ở Bắc Mỹ sau các mẫu Coelophysis. Mẫu vị thành niên bao gồm một phần xương cánh tay, xương mác, và một mảnh răng. Năm 2005, Ronald S. Tykoski gán chi Dilophosaurus cho một mẫu vật (TMM 43646-140) từ Gold Spring, Arizona, nhưng vào năm 2012, Matthew T. Carrano và các đồng nghiệp phát hiện một số khác biệt chi tiết.
Các loài từng được gán cho chi

Vào năm 1984, Welles kết luận rằng mẫu vật năm 1964 (UCMP 77270) không thuộc chi Dilophosaurus mà thuộc về một chi mới, dựa trên sự khác biệt về cấu trúc sọ, đốt sống và xương đùi. Ông nhận thấy cả hai chi đều có cặp đỉnh, nhưng hình dạng chính xác của mào trên Dilophosaurus vẫn chưa được xác định. Welles qua đời vào năm 1997 trước khi kịp đặt tên cho loài khủng long mới, và quan điểm về hai chi riêng biệt dần bị lãng quên. Năm 1999, nhà cổ sinh vật học nghiệp dư Stephan Pickering công bố tên mới cho Dilophosaurus là 'breedorum' dựa trên mẫu năm 1964, để tôn vinh Breed, người đã hỗ trợ thu thập mẫu vật. Tuy nhiên, cái tên này được coi là nudum nomen, tức danh pháp không hợp lệ. Gay chỉ ra vào năm 2005 rằng không có sự khác biệt rõ rệt giữa D. 'breedorum' và các mẫu D. wetherilli khác. Đến năm 2012, Carrano và cộng sự phát hiện nhiều điểm khác biệt giữa mẫu 1964 và mẫu định danh, nhưng cho rằng những khác biệt này chỉ là sự thay đổi cá thể, không phải loài.
Vào năm 1987, một bộ xương khủng long chân thú gần như hoàn chỉnh (KMV 8701) được phát hiện tại hệ tầng Lục Phong ở tỉnh Vân Nam, Trung Quốc. Bộ xương này tương tự như Dilophosaurus, với cặp mào và khe hở giữa xương hàm trên và xương hàm dưới, nhưng có một số điểm khác biệt. Nhà cổ sinh vật học Trung Quốc Shaojin Hu cho rằng đây là một loài mới thuộc chi Dilophosaurus, đặt tên là D. sinensis vào năm 1993 (tiếng Hy Lạp Sinai nghĩa là Trung Quốc). Năm 1998, Matthew C. Lamanna và các đồng nghiệp phát hiện mẫu hóa thạch D. sinensis giống hệt Sinosaurus triassicus, một loài khủng long chân thú từ cùng thành hệ, được đặt tên vào năm 1940. Phát hiện này được xác nhận bởi Lida Xing và các đồng nghiệp vào năm 2013, và mặc dù Guo-Fu Wang và cộng sự xác định loài này thuộc chi Sinosaurus vào năm 2017, họ cho rằng rất có thể đây là loài S. sinensis.
Miêu tả
Dilophosaurus là một trong những loài khủng long săn mồi lớn xuất hiện sớm nhất, thuộc nhóm theropod cỡ trung bình, dù vẫn nhỏ hơn so với những loài khủng long ăn thịt sau này. Kích thước của nó gần tương đương với một con gấu nâu, với cơ thể thon gọn và nhẹ. Mẫu lớn nhất được biết nặng khoảng 400 kg, dài khoảng 7 mét, và sọ dài 590 mm. Mẫu định danh nhỏ hơn, nặng khoảng 283 kg, dài 6,03 mét, với chiều cao hông khoảng 1,36 mét, và sọ dài 523 mm. Dựa trên dấu vết của một loài theropod tương tự như Dilophosaurus và Liliensternus phát hiện vào năm 1996, một số nhà nghiên cứu cho rằng có thể thấy dấu vết của lông vũ quanh bụng và chân. Tuy nhiên, các nhà nghiên cứu khác giải thích hiện tượng này là sự cô đặc trầm tích khi khủng long di chuyển, nhưng vẫn không hoàn toàn bác bỏ khả năng khủng long có lông vũ.
Hộp sọ






Hộp sọ của Dilophosaurus khá lớn so với các xương khác nhưng lại rất mỏng manh. Mõm hẹp từ phía trước, dần thu hẹp về phía đầu tròn. Xương trước-hàm trên dài và thấp khi nhìn từ bên, hình củ hành phồng to khi nhìn từ phía trước, và mặt ngoài của nó ít lõm dần từ mõm đến lỗ mũi. Lỗ mũi nằm xa về phía sau so với các khủng long chân thú khác. Xương trước-hàm gắn rất gần nhau, mặc dù chỉ có một điểm nối chính giữa với xương hàm trên mà không có điểm nối bên, nhưng vẫn tạo thành khớp chắc chắn nhờ sự khóa liên động giữa các mỏm. Mép dưới của xương trước-hàm trên và xương hàm trên tạo ra một khoảng trống gọi là kẽ hở dưới lỗ mũi, tương tự như ở các loài khủng long coelophysoid. Kẽ hở này tạo ra một chỗ trống sâu sau hàng răng của xương trước-hàm trên, gọi là hố dưới lỗ mũi, được bao quanh bởi sống xương hướng xuống của xương-trước hàm trên.
Bề mặt ngoài của xương trước-mõm trên có các lỗ với đủ kích cỡ. Phần trên của hai mỏm mọc chĩa về phía sau hướng thấp rất dài, tạo thành phần lớn viền trên của lỗ mũi dài. Phần này có một độ dốc về phía mặt tiền, làm cho khu vực dưới nó bị lõm. Mặt dưới của đầu mõm trên chứa các ổ cắm răng hình bầu dục. Xương mõm trên nông cạn, biến dạng xung quanh cửa sổ trước hố mắt (một lỗ lớn ở phía trước của mắt), tạo thành một lõm tròn về phía trước và mượt hơn so với phần còn lại của hàm trên. Một lỗ gọi là vùng cửa sổ trước hố mắt mở ra trong rãnh này ở phía trước uốn cong. Lỗ lớn chạy ở phía bên của hàm trên, trên phần lỗ chân răng. Một rãnh dinh dưỡng sâu chạy ngược từ hố dưới lỗ mũi theo đáy của các tấm kẽ răng của hàm trên.
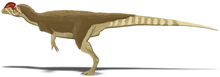
Dilophosaurus sở hữu một cặp mào cao, mỏng và cong (hoặc hình đĩa) dọc theo đỉnh hộp sọ. Các mào chủ yếu được cấu thành từ xương lệ và một phần từ xương mũi. Cặp mào này mở rộng lên trên và giống như một con đà điểu đầu mào với hai đỉnh. Đặc biệt, mỗi đỉnh còn có một phần mở rộng giống như ngón tay. Bề mặt trên của xương mũi giữa các đỉnh khá lõm, và phần mũi của đỉnh phủ lên phần xương lệ. Vì chỉ có một mẫu duy nhất bảo tồn hình dạng của các đỉnh, chưa rõ liệu mào có sự khác biệt giữa các cá thể hay không. Xương lệ có một vành dày ở trên tạo thành biên giới phía trên của cửa sổ trước hố mắt. Xương trước trán tạo thành mái của hố mắt và có một thanh chữ L làm cho bề mặt trên của hố mắt bị lõm. Hố mắt có hình bầu dục và hẹp về phía dưới. Xương gò má có hai mỏm hướng lên trên, mỏm đầu tiên tạo thành phần lề dưới của cửa sổ trước hố mắt, và một phần của lề dưới hố mắt. Xương vuông chiếu vào cửa sổ phía sau mắt tạo thành chu vi hình quả thận. Lỗ chẩm, lớn và hình trái tim, chiếm khoảng một nửa bề rộng của lồi cầu xương chẩm, có một cổ ngắn và một rãnh ở bên.
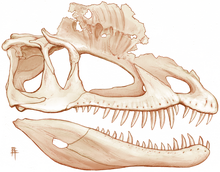
Hàm dưới của Dilophosaurus có vẻ mảnh khảnh và tinh tế ở phía trước, nhưng vùng khớp với hộp sọ rất lớn và nằm sâu quanh cửa sổ hàm dưới. Các cửa sổ này nhỏ hơn so với các loài coelophysoid. Mỏm dưới của cung gò má của hàm dưới dài, và xương surangular ngang. Xương răng phía trước của hàm dưới cong nhẹ chứ không nhọn. Cằm có một lỗ lớn ở mũi và một hàng lỗ nhỏ song song với mép trên của xương răng. Phía bên trong, đường chỉ giữa hàm dưới phẳng và mịn, không có dấu hiệu hợp nhất với nửa đối diện. Một lỗ Meckelian chạy dọc theo bên ngoài của hàm răng.
Dilophosaurus có bốn răng trên mỗi xương đầu mõm của hàm trên, mười hai trên mỗi xương hàm trên, và mười bảy trên mỗi xương răng. Răng dài, thon và cong, với đáy nhỏ. Chúng nén ngang, hình bầu dục ở đáy, dạng thấu kính ở trên, và hơi lõm ở mặt ngoài và trong. Răng lớn nhất của hàm trên thuộc xương ổ răng thứ tư, và chiều cao của các răng giảm dần về phía sau. Răng đầu tiên của hàm trên chỉ hơi hướng về phía trước vì đường viền dưới của mỏm xương đầu mõm của hàm trên hướng thẳng lên. Răng của xương răng nhỏ hơn so với hàm trên. Răng thứ ba hoặc thứ tư và một số coelophysoids là lớn nhất, phù hợp với khoảng cách dưới lỗ mũi của hàm trên. Hầu hết các răng có răng cưa ở các cạnh trước và sau, được bù bằng các rãnh dọc, và nhỏ hơn ở phía trước. Có từ 31 đến 41 răng cưa ở các cạnh trước, và 29 đến 33 ở cạnh sau. Ít nhất các răng thứ hai và thứ ba có răng cưa, nhưng răng thứ tư thì không. Răng được phủ bởi lớp men mỏng, dày từ 0,1 đến 0,15 mm, kéo dài xa về phía chân của chúng. Các ổ răng hình elip đến gần tròn, lớn hơn các chân răng, có thể là răng đã được giữ lỏng lẻo trong hàm. Mặc dù số lượng ổ răng dường như cho thấy răng đông đúc, chúng khá xa nhau do kích thước lớn của ổ. Hàm có nhiều răng thay thế ở các giai đoạn mọc khác nhau. Các tấm kẽ răng rất thấp.
Khung xương sau sọ
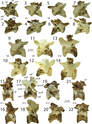


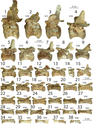
Dilophosaurus có 10 đốt sống cổ, 14 đốt sống lưng và 45 đốt sống đuôi. Cái cổ dài của nó có thể cong gần 90° nhờ vào cấu trúc của hộp sọ và vai, giữ cho hộp sọ ở tư thế nằm ngang. Các đốt sống cổ của nó rất nhẹ, với thân đốt sống được làm rỗng nhờ các khoang pleurocoel và centrocoel. Vòm của các đốt sống cổ có chonoses lớn, tạo ra các rãnh hình nón khiến xương trở nên rất mỏng manh. Các thân đốt sống cổ có hình phẳng-lõm, phẳng ở phía trước và lõm ở phía sau, tương tự như ở loài Ceratosaurus. Điều này cho thấy cổ của nó rất linh hoạt, dù các sườn cổ dài và chồng lên nhau đã hợp nhất với thân đốt sống cổ. Xương sườn cổ mảnh mai và có thể uốn cong dễ dàng. Đốt atlas (đốt sống cổ thứ nhất) có một khối thân đốt nhỏ và có sự lõm ở phía trước, tạo thành một cái cúp cho lồi cầu xương chẩm ở mặt sau của hộp sọ. Đốt trục (đốt sống cổ thứ hai) có cột sống nặng và các mỏm khớp dưới nối với các mỏm khớp trên dài cong của đốt sống cổ thứ ba. Các centra và mỏm gai của đốt sống cổ dài và thấp, với mỏm gai tạo thành hình chữ thập Mantit khi nhìn từ trên xuống, một đặc điểm đặc trưng của loài này. Các xương sống thần kinh của đốt sống lưng cũng thấp và mở rộng ra trước và sau, tạo thành các điểm bám chắc chắn cho dây chằng. Các đốt sống xương cùng, chiếm chiều dài của lưỡi xương chậu, dường như không hợp nhất. Các centra của đốt sống đuôi đều có chiều dài tương tự nhau, nhưng đường kính nhỏ dần về phía sau, với tiết diện từ hình elip đến hình tròn.
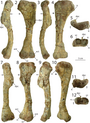
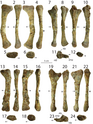




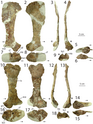
Các xương vai có chiều dài trung bình và lõm vào bên trong để phù hợp với độ cong của cơ thể. Xương vai khá rộng, đặc biệt là phần trên có hình chữ nhật, đây là một đặc điểm độc đáo. Xương quạ có hình elip và không hợp nhất với xương vai. Cánh tay rất mạnh mẽ, có hố sâu và các mõm lớn để tạo diện tích cho các cơ và dây chằng bám vào. Xương cánh tay (xương chi cánh tay trên) lớn và mảnh khảnh, với một cục xương cẳng tay lớn, và xương trụ (xương chi cánh tay dưới) to và thẳng, với một mỏm khủyu lớn. Bàn tay có bốn ngón: ngón đầu tiên ngắn hơn nhưng mạnh hơn hai ngón sau, với móng vuốt lớn, và hai ngón sau dài hơn và mảnh hơn, với móng vuốt nhỏ hơn. Ngón thứ ba có kích cỡ nhỏ hơn, và ngón thứ tư là di tích (còn lại nhưng không có chức năng). Đỉnh của xương chậu cao nhất ở cuống xương chậu (mõm chĩa xuống của xương chậu), và phần bên ngoài của nó là lõm. Chân của xương mu chỉ hơi mở rộng, trong khi đầu thấp hơn được mở rộng nhiều hơn với đốt háng, và có một trục rất mỏng. Chân sau lớn, với xương đùi (xương chân trên) dài hơn xương chày (xương chân dưới), trái ngược với loài Coelophysis. Xương đùi to, với trục hình cong chữ S, và đốt chuyển lớn nằm trên trục. Xương chày có mấu lồi phát triển và mở rộng ở đầu thấp. Xương sên (xương mắt cá chân) tách rời khỏi xương chày và xương gót chân, hình thành một nửa lỗ cho xương mác. Loài này có đôi chân dài và to với ba ngón chân phát triển tốt mang các móng vuốt lớn. Ngón chân thứ ba là mạnh nhất, và ngón chân cái nhỏ được giữ cách xa mặt đất.
Phân loại

Năm 1954, Welles đã xác định Dilophosaurus là thành viên của họ megalosaur, nhưng sau đó đã thay đổi quan điểm vào năm 1970 khi phát hiện chúng có mào. Đến năm 1974, Welles và nhà cổ sinh vật học người Mỹ Robert A. Long đã phát hiện rằng Dilophosaurus thuộc về nhóm khủng long đầu gồ. Vào năm 1984, Welles ghi nhận rằng Dilophosaurus có các đặc điểm của cả khủng long đuôi rỗng và Carnosaurus, hai phân nhóm chính của khủng long chân thú, dựa trên kích thước cơ thể. Ông phát hiện Dilophosaurus có mối quan hệ gần gũi với các loài khủng long thuộc họ Coelophysoidea, đặc biệt là chi Liliensternus.
Năm 1988, nhà cổ sinh vật học người Mỹ Gregory S. Paul phân loại các halticosaur vào phân nhóm của họ Coelophysidae, và cho rằng Dilophosaurus có thể là hậu duệ trực tiếp của Coelophysis. Paul cũng xem xét khả năng các đại long xương gai chính là hậu duệ của các dilophosaur, dựa trên các điểm tương đồng như kẽ hở hàm trên, vị trí lỗ mũi và các răng thon của loài Baryonyx. Vào năm 1994, nhà cổ sinh vật học người Mỹ Thomas R. Holtz đã xếp Dilophosaurus vào nhóm Coelophysoidea, nhưng tách biệt khỏi Coelophysidae. Ông xếp Coelophysoidea vào nhóm Ceratosauria. Đến năm 2000, nhà cổ sinh vật học người Mỹ James H. Madsen và Welles đã chia tách Ceratosauria thành họ Ceratosauridae và họ Khủng long hai mào (trong đó Dilophosaurus là thành viên duy nhất).

Năm 1998, Lamanna và các cộng sự đã chỉ ra rằng sau khi Dilophosaurus được phát hiện với đôi mào, nhiều loài khủng long có mào tương tự cũng được phát hiện (bao gồm cả Sinosaurus), và do đó đặc điểm này không phải là đặc hữu và không nên quá lạm dụng trong các nghiên cứu phân loại. Năm 2005, nhà cổ sinh vật học người Úc Adam M. Yates đã mô tả chi Dracovenator mới phát hiện ở Nam Phi, và cho rằng nó có mối quan hệ gần gũi với Dilophosaurus và Zupaysaurus. Phân tích nhánh học của ông cho thấy chúng không thuộc nhóm Coelophysoidea, mà thuộc nhóm Neotheropoda 'tân tiến' hơn. Ông đề xuất rằng nếu Dilophosaurus thực sự tiến hóa hơn nhóm Coelophysoidea, thì các đặc điểm tương đồng của nó với nhóm này có thể là thừa hưởng từ tổ tiên khủng long chân thú 'cơ bản' hơn, cho thấy rằng mọi loài khủng long chân thú đều đã trải qua 'giai đoạn coelophysoid' trong quá trình tiến hóa của chúng.
Năm 2007, nhà cổ sinh vật học người Mỹ Nathan D. Smith và các cộng sự phát hiện hóa thạch của chi Cryolophosaurus, có mối quan hệ gần gũi với Dilophosaurus, và nhóm này cũng liên quan đến Dracovenator và Sinosaurus. Nhóm này tiến hóa hơn Coelophysoidea nhưng vẫn cơ bản hơn Ceratosauria, dẫn đến phân loại của các khủng long chân thú cơ bản có dạng bậc thang. Đến năm 2012, Carrano và các đồng nghiệp chỉ trích phân loại của Smith vì chỉ dựa vào đặc điểm mào mà không chú ý đến chi tiết xương sau sọ. Họ cho rằng Dilophosaurus thuộc nhóm coelophysoid, với Cryolophosaurus và Sinosaurus là các thành viên tiên tiến hơn của nhóm Tetanurae.
Nhà cổ sinh vật học người Bỉ Kouthe Hendrickx và các đồng nghiệp đã định nghĩa nhóm Dilophosauridae vào năm 2015, bao gồm Dilophosaurus và Dracovenator. Họ lưu ý rằng mặc dù sự phân loại này vẫn còn đang được xem xét, nhóm này có vẻ tiến hóa hơn nhiều so với Coelophysoidea và là nhóm chị em của Averostra. Dilophosauridae có nhiều đặc điểm chung với Coelophysoidea, như kẽ hở dưới mũi và răng trước của xương hàm trên hướng về phía trước, đồng thời cũng có những đặc điểm giống Averostra như cửa sổ trước của xương hàm trên và số lượng răng hàm trên giảm. Họ cho rằng các đỉnh mào của Cryolophosaurus và Sinosaurus có thể là kết quả của sự tiến hóa hội tụ hoặc di truyền từ tổ tiên chung. Sơ đồ phát sinh chủng loài dưới đây dựa trên các đề xuất của Hendrickx và đồng nghiệp cũng như các nghiên cứu trước đó.
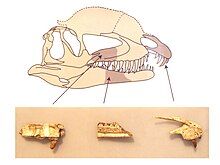
| Neotheropoda |
| |||||||||||||||||||||||||||||||||||||||||||||||||||
Dấu vết hóa thạch
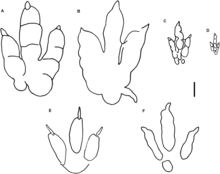
Nhiều đơn vị phân loại dấu chân (ichnotaxa) đã được gán cho chi Dilophosaurus hoặc các khủng long chân thú tương tự. Vào năm 1971, Welles đã ghi nhận nhiều dấu chân khủng long ở các tầng đá cách mặt đất 14 mét (45 ft) và 112 mét (367 ft) của thành hệ Kayenta, ngay dưới nơi tìm thấy mẫu định danh Dilophosaurus. Các dấu chân ở tầng thấp thuộc loại tridactyl (ba ngón) và có thể thuộc về Dilophosaurus; Welles đã đặt tên cho chi dấu chân là Dilophosauripus williamsi để vinh danh Williams, người đã phát hiện bộ xương Dilophosaurus đầu tiên. Mẫu chuẩn của loài dấu chân là bản in dấu chân mã số UCMP 79690-4, cùng với ba bản cast dấu chân khác trong nhóm mẫu gốc. Đến năm 1984, Welles thừa nhận không thể xác minh chắc chắn rằng đây là dấu chân của Dilophosaurus. Năm 1996, Michael Morales và Scott Bulkey báo cáo một đường dấu chân thuộc chi Eubrontes từ hệ tầng Kayenta, do một khủng long chân thú lớn để lại. Họ cho rằng dấu vết này có thể không phải của Dilophosaurus vì kích thước của con vật tạo ra dấu chân lớn hơn nhiều so với kích thước của Dilophosaurus.
Nhà cổ sinh vật học người Ba Lan Gerard Gierliński đã nghiên cứu dấu chân tridactyl từ dãy núi Świętokrzyskie ở Ba Lan và vào năm 1991 kết luận rằng chúng thuộc về một loài khủng long chân thú tương tự như Dilophosaurus. Ông đã đặt tên cho dấu chân là Grallator (Eubrontes) soltykovensis, với mẫu cast dấu chân MGIW 1560.11.12 trở thành mẫu định danh. Năm 1994, Gierliński ấn định dấu chân từ năm 1974 ở Thành hệ Höganäs, Thụy Điển là loài G. (E.) soltykovensis. Năm 1996, ông đã gán dấu vết AC 1/7 từ Thành hệ Thác nước Turners, Massachusetts cho một loài khủng long chân thú giống Dilophosaurus và Liliensternus, và đặt tên cho nó là Grallator minisculus. Nhà cổ sinh vật học người Séc Martin Kundrát đồng ý rằng có thể thấy dấu vết lông vũ trong dấu vết này vào năm 2004, nhưng một số nhà nghiên cứu khác như Martin Lockley (2003) và Anthony J. Martin (2004) cho rằng những vết hằn đó chỉ là di vật trầm tích. Martin và các đồng nghiệp cũng đã kiểm định lại dấu vết này và phân loại nó là Fulicopus lyellii.
Nhà cổ sinh vật học người Mỹ Robert E. Weems (2003) cho rằng dấu vết Eubrontes không phải do một khủng long chân thú tạo ra mà do một khủng long dạng chân thằn lằn giống Plateosaurus. Weems cho rằng đơn vị phân loại dấu chân Kayentapus hopii, do Welles đặt tên vào năm 1971, là dấu chân phù hợp nhất với Dilophosaurus. Kết luận này dựa chủ yếu vào góc rộng giữa ngón ba và bốn, và ngón thứ tư của bàn chân nguyên mẫu cũng có góc tương tự. Cùng năm đó, Emma Rainforth phản biện rằng sự tỏa ra của ngón chân nguyên mẫu chỉ là biến dạng cá thể, và khẳng định Eubrontes là ứng cử viên hợp lý hơn cho dấu chân của Dilophosaurus.
Cổ sinh học
Di chuyển

Welles tưởng tượng Dilophosaurus như một sinh vật năng động, di chuyển bằng hai chân như một con đà điểu khổng lồ. Ông cho rằng các chi trước là vũ khí mạnh mẽ, linh hoạt và không được sử dụng để di chuyển. Ông nhận xét rằng hai bàn tay có khả năng nắm và chém, có thể giao nhau và nâng lên khoảng hai phần ba chiều cao của cổ. Welles giả định rằng trong tư thế ngồi, con vật sẽ dựa vào 'bàn chân' lớn của đốt háng, cùng với đuôi và chân. Vào năm 1990, các nhà cổ sinh vật học Stephen và Sylvia Czerkas cho rằng xương chậu yếu của Dilophosaurus có thể là sự thích nghi cho môi trường nước, hỗ trợ trọng lượng của chúng, và chúng có thể là loài bơi lội hiệu quả. Tuy nhiên, họ nghi ngờ rằng chúng bị hạn chế hoàn toàn trong môi trường nước, do chân sau mạnh mẽ và tỷ lệ cơ thể cho phép chúng di chuyển nhanh nhẹn và dẻo dai trên cạn.
Vào năm 2005, các nhà cổ sinh vật học Phil Senter và James H. Robins đã nghiên cứu khả năng di chuyển của các chi trước của Dilophosaurus và các khủng long chân thú khác. Họ phát hiện rằng Dilophosaurus có thể kéo xương cánh tay về phía sau gần như song song với xương bả vai, nhưng không thể di chuyển nó về phía trước theo hướng thẳng đứng. Khuỷu tay có thể mở ra hoàn toàn và uốn cong một góc vuông, nhưng không đạt được góc hoàn toàn. Ngón tay có vẻ không thể chủ động kéo dài ra phía sau quá mức bình thường, nhưng có thể bị tác động thụ động để tránh sự trật khớp trong các chuyển động dữ dội. Một nghiên cứu năm 2015 của Senter và Robins đưa ra các khuyến nghị về cách tái tạo tư thế chân trước của khủng long hai chân, dựa trên nhiều cuộc kiểm tra loài khác nhau, bao gồm cả Dilophosaurus. Xương bả vai được giữ ngang, hướng nghỉ ngơi của khuỷu tay gần góc vuông, và hướng của bàn tay không lệch nhiều so với cánh tay dưới.

Các dấu chân Dilophosauripus được Welles báo cáo vào năm 1971 đều ở cùng một tầng cao và được mô tả như là 'dấu vết của sân gà', với một số ít tạo thành một lối đi. Các dấu chân in chìm trong bùn, cho phép chân chìm xuống 5–10 cm. Các dấu chân rất không đồng đều, và độ rộng khác nhau của các dấu chân ngón chân cho thấy bùn đã bám vào chân. Các vết hằn khác nhau tùy thuộc vào chất nền và cách tạo ra chúng; đôi khi chân đặt xuống trực tiếp, nhưng thường có trượt ngược hoặc xuôi khi bàn chân chạm xuống. Vị trí và góc của các ngón chân thay đổi đáng kể, cho thấy chúng rất linh hoạt. Dấu chân Dilophosauripus có ngón chân thứ hai với đế dày và móng vuốt thẳng, dài, và thẳng hàng với trục của miếng đệm ngón chân. Một dấu chân thiếu móng vuốt thứ hai, có thể do chấn thương. Vào năm 1984, Welles giải thích việc ba cá thể được tìm thấy gần nhau và các dấu chân chéo gần đó như dấu hiệu cho thấy Dilophosaurus đi theo bầy. Gay đồng ý rằng chúng có thể đã di chuyển theo nhóm nhỏ, nhưng lưu ý rằng không có bằng chứng trực tiếp cho tập tính này và lũ quét có thể đã phân tán xương từ các cá thể khác nhau và đưa chúng lại gần nhau.
Vào năm 2009, Milner và nhóm của ông đã khảo sát đường dấu chân Utah SGDS.18.T1, bao gồm các dấu chân với đuôi kéo lê và dấu vết nghỉ ngơi bất thường hơn trong sa thạch của hồ. Đường dấu chân bắt đầu với con vật di chuyển song song với bờ hồ, sau đó dừng lại bên một gờ với cả hai chân song song. Nó hạ thân mình xuống, đưa bàn chân và mông tại khung chậu xuống đất, tạo ra những vết hằn 'gót chân' đối xứng và dấu vết tròn của khung chậu. Phần đuôi gần cơ thể giữ khỏi mặt đất, còn phần đuôi xa hơn tiếp xúc với mặt đất. Con vật nằm trên một dốc cho phép nó đưa cả hai tay xuống gần bàn chân. Sau khi nghỉ, con khủng long tiến về phía trước, để lại dấu vết mới với bàn chân và khung chậu, nhưng không có dấu tay. Bàn chân phải bước lên dấu của bàn tay phải, và móng thứ hai của bàn chân trái tạo ra một vết lê từ vị trí nghỉ đầu tiên đến vị trí tiếp theo. Sau một thời gian, con vật đứng dậy và di chuyển về phía trước, với chân trái trước, và sau khi duỗi chân thẳng, nó đi qua phần còn lại của bề mặt, để lại những vết kéo mỏng bởi phần cuối đuôi.
Hành vi cúi là rất hiếm gặp ở khủng long chân thú, và SGDS.18.T1 là đường dấu chân duy nhất có vết in rõ ràng về bàn tay của chúng, cung cấp thông tin quý giá về cách sử dụng chi trước. Tư thế cúi khá giống các loài chim hiện đại, cho thấy khủng long chân thú ban đầu giữ hai lòng bàn tay đối diện nhau, hướng vào trong. Điều này cho thấy tư thế như vậy xuất hiện sớm trong dòng dõi khủng long chân thú và có thể là đặc điểm chung của tất cả khủng long chân thú. Khủng long chân thú thường được mô tả với lòng bàn tay hướng xuống dưới, nhưng các nghiên cứu về giải phẫu chức năng cho thấy chúng, như chim, không thể quay sấp hay lật ngửa cánh tay. Đường dấu chân cho thấy các chân sau giữ đối xứng, với trọng lượng cơ thể phân bổ đều giữa các bàn chân, đặc điểm này cũng thấy ở các loài chim Ratite. Milner và các đồng nghiệp cũng bác bỏ ý kiến cho rằng đường vết Kayentapus minor được Weems báo cáo cho thấy dấu ấn lòng bàn tay của khủng long chân thú đi bộ tứ chi. Weems cho rằng con vật có thể đi bằng bốn chi khi di chuyển chậm và lòng bàn tay liên tục chạm đất. Milner và các đồng nghiệp cho rằng đề xuất này không cần thiết, và thay vào đó, đường dấu này được thực hiện giống như ở SGDS.18.T1 nhưng không để lại dấu vết của các ngón tay.
Chế độ ăn
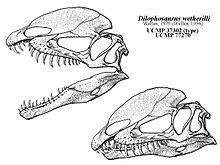
Welles phát hiện rằng Dilophosaurus không có vết cắn mạnh do khoảng trống dưới xương mũi. Ông cho rằng chúng sử dụng răng trước hàm để nhổ và xé, không phải cắn, và răng hàm trên xa về sau hơn dùng để đâm và cắt. Ông nghĩ rằng chúng có thể là loài ăn xác thay vì kẻ săn mồi, và nếu thực sự giết mồi, chúng sẽ dùng tay thay vì hàm. Welles không tìm thấy bằng chứng sọ di trong hộp sọ của Dilophosaurus, một đặc điểm cho phép các xương hộp sọ di chuyển tương đối với nhau.
Năm 1986, nhà cổ sinh vật học Robert T. Bakker từ Mỹ phát hiện Dilophosaurus với cổ và hộp sọ khổng lồ, cùng với hàm răng trên lớn hơn. Điều này cho thấy loài này có thể đã phát triển để tiêu diệt con mồi lớn và đủ mạnh để tấn công các động vật ăn cỏ từ kỷ Jura sớm. Tuy nhiên, năm 1988, Paul đã phản bác quan điểm cho rằng Dilophosaurus là một kẻ ăn thịt và cho rằng đây là một nhận định sai. Ông lập luận rằng mõm của Dilophosaurus không chỉ đáng chú ý mà còn nguy hiểm hơn nhiều so với móng vuốt, và có thể loài này săn bắt nhiều prosauropod hoặc các loài vật nhỏ hơn hiệu quả hơn những loài có kích thước tương tự. Paul cũng mô tả Dilophosaurus nhảy trên đuôi trong khi tấn công kẻ thù, giống như một con chuột túi.
Năm 2005, nghiên cứu về lý thuyết dầm Euler–Bernoulli của nhà nghiên cứu cổ sinh vật học người Canada, François Therrien và cộng sự đã chỉ ra rằng lực cắn của hàm dưới Dilophosaurus giảm dần khi tiến vào trong miệng. Chỉ số này, với cái cằm hếch lên, răng hình 'hoa hồng' và vùng tiếp hợp ghép gia cố (tương tự như spinosaurid), được thiết kế để bắt và giữ con mồi có kích thước nhỏ. Các đặc điểm của hàm dưới của chúng giống như ở mèo và cá sấu, nơi mà hàm được sử dụng để cắn mạnh và khống chế con mồi. Tải trọng lên hàm dưới liên quan đến việc bắt con mồi nhỏ và có thể chúng cắn nhiều phát nhỏ để làm mồi bị thương, rồi sau đó giữ chặt con mồi bằng hàm trước. Con mồi sau đó bị di chuyển vào trong hàm và bị giết bởi các vết cắn mạnh, với hàm răng nén ngang. Các tác giả cho rằng nếu Dilophosaurus ăn con mồi nhỏ, thì kích thước của các bầy săn mồi có thể bị hạn chế.
Năm 2007, Milner và James I. Kirkland đã đề xuất rằng Dilophosaurus có các đặc điểm cho thấy nó có thể ăn cá. Họ chỉ ra rằng phần cuối của hàm dưới mở rộng sang hai bên, tạo thành hình 'hoa hồng' với những chiếc răng lồng vào nhau, tương tự như các loài trong họ spinosaurid, được biết đến với khả năng ăn cá và cá sấu Ấn Độ, thường xuyên ăn cá. Các lỗ mũi ở xa mõm, giống như ở spinosaurid, có lỗ mũi rút lại nhiều hơn, có thể giúp hạn chế nước vào lỗ mũi khi bắt cá. Cả hai nhóm khủng long cũng có cánh tay dài với móng vuốt phát triển tốt, giúp hỗ trợ trong việc bắt cá. Hồ Dixie, một hồ nước lớn kéo dài từ Utah đến Arizona và Nevada, có thể cung cấp lượng cá dồi dào sau sự kiện tuyệt chủng Kỷ Trias–Jura.
Chức năng của mào

Welles thừa nhận rằng các giả thuyết về chức năng của các đỉnh của Dilophosaurus chủ yếu chỉ là những phỏng đoán. Mặc dù các đỉnh không có rãnh để chứa mạch máu, ông vẫn cho rằng chúng có thể đã được dùng để điều chỉnh nhiệt độ cơ thể. Ông cũng gợi ý rằng các đỉnh có thể được sử dụng để nhận diện đồng loại hoặc để trang trí. Czerkas cho rằng các mào không thể được dùng trong các cuộc chiến do cấu trúc mỏng manh của chúng dễ bị phá hủy. Họ cho rằng chúng có thể là một dấu hiệu trực quan để thu hút bạn tình, và thậm chí giúp điều chỉnh nhiệt độ. Vào năm 1990, nhà cổ sinh vật học người Mỹ Walter P. Coombs cho rằng các mào có thể đã được tô điểm bằng màu sắc để các con đực thể hiện sự quyến rũ.
Vào năm 2011, các nhà cổ sinh vật học người Mỹ Kevin Padian và John R. Horner đã đưa ra giả thuyết rằng 'các cấu trúc kỳ lạ' trên khủng long (bao gồm cả mào, diềm, sừng và vòm) chủ yếu được dùng để nhận diện loài, bác bỏ các giả thuyết khác do thiếu bằng chứng rõ ràng. Họ lưu ý rằng có quá ít mẫu vật của các loài khủng long chân thú với các bộ phận trưng bày, bao gồm Dilophosaurus, để các nhà khoa học có thể kiểm tra chức năng tiến hóa của mào qua phương pháp thống kê, và xem liệu chúng có liên quan đến lưỡng hình tình dục hoặc trưởng thành tình dục hay không.
Cổ bệnh lí học
Welles đã phát hiện nhiều loại bệnh lý cổ xưa, chẳng hạn như chấn thương và dị tật, trên Dilophosaurus. Mẫu định danh có một rãnh trên cung đốt sống của một đốt sống cổ, có thể là dấu hiệu của chấn thương hoặc bị dập nát, và hai hố trên xương cánh tay phải có thể là dấu hiệu của áp-xe (tập hợp mủ). Welles cũng chỉ ra rằng xương cánh tay bên trái nhỏ hơn và mảnh mai hơn bên phải, nhưng điều này ngược lại ở cẳng tay. Vào năm 2001, nhà cổ sinh vật học người Úc Ralph Molnar cho rằng điều này là do sự bất đối xứng dao động trong quá trình phát triển. Đặc điểm này có thể xuất phát từ sự căng thẳng trong quần thể động vật, ví dụ do sự xáo trộn trong môi trường sống, và có thể cho thấy áp lực chọn lọc tự nhiên mạnh mẽ hơn. Sự bất đối xứng cũng có thể do các chấn thương trong giai đoạn phát triển ban đầu của loài, với sự phân phối ngẫu nhiên theo thời gian. Một nghiên cứu năm 2001 của nhà cổ sinh vật học người Mỹ Bruce Rothschild và các đồng nghiệp đã kiểm tra 60 xương bàn chân Dilophosaurus để tìm dấu hiệu gãy xương do áp lực (do các hoạt động tạo áp lực lặp đi lặp lại), nhưng không phát hiện điều gì bất thường. Chấn thương như vậy có thể do lối sống săn mồi rất tích cực.
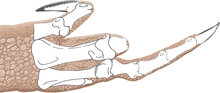
Năm 2016, Senter và Sara L. Juengst đã kiểm tra các nghiên cứu về mẫu định danh và phát hiện rằng nó chứa số lượng lớn nhất và đa dạng nhất các bệnh lý ác tính trên dầm ngực và chân trước so với bất kỳ loài khủng long chân thú nào đã được mô tả. Một số chấn thương là độc nhất cho loài này. Chỉ có sáu loài khủng long ăn thịt khác được biết đến với nhiều hơn một loại bệnh lý trên đai ngực và chân trước. Mẫu vật gốc có tám xương bị ảnh hưởng, trong khi không có mẫu vật nào khác của khủng long chân thú có hơn bốn xương bị ảnh hưởng. Bên trái của con vật có xương vai và xương quay bị gãy, cùng với áp-xe ở xương trụ và xương đốt ngón ngoài của ngón tay cái. Bên phải có xương bị xoắn quanh trục cánh tay, ba khối u xương trên xương quay, một bề mặt khớp bị rời trên xương bàn tay thứ ba, và biến dạng trên xương đốt ngón đầu tiên của ngón thứ ba. Ngón tay này đã bị biến dạng vĩnh viễn và không thể cử động uốn. Các biến dạng của cánh tay và ngón tay thứ ba có thể do chứng loạn sản xương, điều này chưa được báo cáo từ khủng long phi chim trước đây nhưng đã được biết đến ở các loài chim. Bệnh này có thể gây đau ở một chi, khiến chim phải sử dụng các chi còn lại, dẫn đến xoắn.
Số lượng các sự kiện chấn thương dẫn đến những dấu hiệu này không rõ ràng, và có thể tất cả các bệnh lý đều do một sự kiện duy nhất gây ra, chẳng hạn như va chạm với cây hoặc đá trong khi chiến đấu với một con vật khác, có thể gây ra vết thương đâm thủng do móng vuốt. Vì tất cả các vết thương đã lành, chắc chắn rằng con Dilophosaurus này đã sống sót qua nhiều tháng, có thể là nhiều năm. Trong thời gian vết thương lành, chi trước của nó có thể không thể bắt mồi, do đó nó có thể đã phải nhịn ăn dài hoặc săn những con mồi đủ nhỏ để có thể bắt bằng miệng và chân hoặc một chi trước. Theo Senter và Juengst, mức độ đau đớn mà nó có thể đã trải qua cho thấy nó là một con vật khỏe mạnh. Họ lưu ý rằng các cổ bệnh lý ở khủng long chưa được báo cáo đầy đủ, và dù Welles đã mô tả chi tiết về mẫu gốc, ông chỉ đề cập đến một trong những bệnh lý được phát hiện. Họ cho rằng các bệnh lý như vậy thường bị bỏ qua vì chúng được coi là đặc điểm của loài hơn là sự bất thường hoặc vì các triệu chứng rất khó nhận ra.
Cổ sinh thái

Dilophosaurus xuất hiện từ hệ tầng Kayenta, nằm trong giai đoạn Sinemurian và Pliensbachian của kỷ Jura sớm, khoảng 196-183 triệu năm trước. Hệ tầng Kayenta thuộc nhóm Glen Canyon, bao gồm các thành hệ ở phía bắc Arizona, đông nam Utah, tây Colorado và tây bắc New Mexico. Hệ tầng này chủ yếu bao gồm hai loại đá: một là đá bột kết và một là sa thạch. Đá bột kết phân bố rộng rãi ở Arizona, trong khi đá sa thạch có mặt ở phía bắc Arizona, miền nam Utah, tây Colorado và tây bắc New Mexico. Hệ tầng này chủ yếu được lắng đọng bởi các dòng sông, với các tướng đá phù sa là phần lắng đọng chậm hơn của hệ thống sông. Sự lắng đọng kết thúc ở cồn lấn chiếm đã trở thành sa thạch Navajo. Niên đại xác định bằng phép đo phóng xạ cho hệ tầng này chưa được thực hiện và mối tương quan địa tầng dựa trên phương pháp đo phóng xạ từ hóa thạch động vật có xương sống, từ địa tầng và bằng chứng của phấn hoa.
Hệ tầng Kayenta đã cung cấp một bộ sưu tập phong phú và đang ngày càng phong phú hơn của nhiều loại sinh vật. Hóa thạch chủ yếu được tìm thấy trong các tướng đá bột kết. Động vật không có xương sống bao gồm vi sinh vật hoặc 'tảo', gỗ hóa đá, dấu vết thực vật, hai mảnh vỏ và ốc sên nước ngọt, ostracod, và hóa thạch dấu vết của sinh vật không xương sống. Động vật có xương sống được biết đến từ cả hóa thạch cơ thể và hóa thạch dấu vết. Các hóa thạch cơ thể bao gồm cá mập hybodont, cá xương không xác định, cá phổi, kỳ nhông, ếch Prosalirus, caecilia Eocaecilia, rùa Kayentachelys, nhiều loài crocodylomorph như Calsoyasuchus, Eopneumatosuchus, Kayentasuchus và Protosuchus, và loài dực long Rhamphinion. Ngoài Dilophosaurus, còn nhiều loài khủng long khác được biết đến như khủng long chân thú Megapnosaurus (có thể có mào và cần một tên chi mới), và Kayentavenator, khủng long dạng chân thằn lằn Sarahsaurus, cùng nhiều loài khủng long bọc giáp như Scelidosaurus và Scutellosaurus. Bộ Một cung bên bao gồm tritylodontid Dinnebitodon, Kayentatherium, và Oligokyphus, morganucodontid, động vật có vú sớm Dinnetherium, và một động vật có vú haramiyid. Phần lớn các phát hiện này đến từ vùng lân cận Gold Spring, Arizona. Hóa thạch dấu vết của động vật có xương sống bao gồm phân hóa thạch và dấu chân của các Cung thú, thú có vú giống thằn lằn và nhiều loại khủng long.
Liên kết ngoài
- “Dilophosaurus!”. Một triển lãm được thuyết minh 'hướng dẫn'. Thuyết minh bởi Samuel P. Welles.Quản lý CS1: khác (liên kết)
- Giới thiệu về Dilophosaurus
- Các lỗi trong Jurassic Park Lưu trữ 2007-12-19 tại Wayback Machine




















Theropoda |
|---|
| Thẻ nhận dạng đơn vị phân loại |
|---|

