
Hệ thống vận chuyển điện tử (tiếng Anh: electron transport chain (ETC)) là quá trình chuyển giao điện tử từ các chất cho điện tử (như NADH) tới các chất nhận điện tử (như oxy), qua đó tạo ra gradient proton qua lớp màng sinh chất. Sự chênh lệch này cung cấp năng lượng cho enzyme ATP synthase để chuyển đổi thành năng lượng hóa học dưới dạng adenosine triphosphate (ATP). Đây là cơ chế tế bào dùng để tạo năng lượng từ ánh sáng trong quang hợp và từ các phản ứng oxy hóa khử như hô hấp tế bào.
Trong lục lạp, ánh sáng phân giải nước thành oxy và NADP chuyển thành NADPH, đồng thời ion H được chuyển qua màng lục lạp. Tại ti thể, oxy chuyển thành nước, NADH thành NAD và succinate thành fumarate, tạo nên gradient proton. Mặc dù một số vi khuẩn thực hiện chuỗi vận chuyển điện tử tương tự như trong ti thể và lạp thể, một số khác dùng các chất nhận và cho điện tử khác. Cả chuỗi vận chuyển điện tử trong quang hợp và hô hấp đều có hiện tượng rò rỉ điện tử sớm tới oxy, tạo superoxide và tăng cường sự căng thẳng oxy hóa.
Nhìn tổng quan
Hệ thống vận chuyển điện tử bao gồm các phản ứng oxy hóa khử xảy ra ở các vị trí khác nhau, trong đó các điện tử (electron) được chuyển từ một phân tử cho điện tử đến một phân tử nhận điện tử. Năng lượng chính điều khiển các phản ứng này là năng lượng tự do Gibbs của các chất phản ứng và sản phẩm. Năng lượng tự do Gibbs là năng lượng khả dụng ('tự do') để thực hiện công việc. Bất kỳ phản ứng nào làm giảm tổng năng lượng tự do Gibbs của hệ thống sẽ tự động xảy ra. Quá trình chuyển điện tử bắt đầu từ một chất cho điện tử đến một chất nhận điện tử.
ATP synthase, một enzyme với cấu trúc được bảo tồn tốt, được cung cấp năng lượng bởi gradient điện hóa proton qua màng, tạo ra từ chuỗi phản ứng oxy hóa khử. Chức năng của hệ thống vận chuyển điện tử là tạo ra gradient điện hóa này. Gradient điện hóa xuyên màng có thể kích thích sự di chuyển các nguyên tử qua màng sinh chất, và có thể kích hoạt các hoạt động cơ học như sự xoay của tiên mao hoặc sản xuất ATP để cung cấp năng lượng cho tế bào. Một lượng nhỏ ATP được sản xuất từ phosphoryl hóa cấp độ chất nền, ví dụ như thủy phân glycogen. Tuy nhiên, phần lớn ATP trong các sinh vật được tạo ra từ hệ thống vận chuyển điện tử, còn một phần nhỏ từ quá trình lên men.
Hệ thống vận chuyển điện tử trong ti thể
Hầu hết các tế bào eukaryote chứa ti thể với chức năng sản xuất ATP thông qua chu trình Krebs, oxy hóa axit béo và oxy hóa amino acid. Tại lớp màng trong của ti thể, các điện tử (electron) từ NADH và succinate di chuyển qua hệ thống vận chuyển điện tử đến oxy, oxy sẽ bị khử thành nước. Hệ thống vận chuyển điện tử bao gồm hơn 15 chất cho và nhận điện tử dạng enzyme. Mỗi chất cho điện tử chuyển điện tử đến các chất nhận có ái lực cao hơn với điện tử, và các chất nhận này tiếp tục chuyển điện tử đến chất nhận tiếp theo; cuối cùng, điện tử được chuyển đến oxy, chất nhận cuối cùng và có ái lực cao nhất trong hệ thống. Quá trình chuyển điện tử này giải phóng năng lượng để tạo ra gradient proton (giữa hai không gian ti thể được ngăn cách bởi màng ti thể) bằng cách bơm proton vào khoảng không giữa các màng ti thể, tạo ra một trạng thái nhiệt động lực học với điện thế để thực hiện các hoạt động. Trong khi oxy hóa kiểu 'đốt cháy' giải phóng phần lớn năng lượng dưới dạng nhiệt, oxy hóa sinh học ở ti thể giải phóng phần lớn năng lượng này dưới dạng có thể sử dụng được nhờ các cơ chế trong hệ thống vận chuyển điện tử.
Tuy nhiên, một số điện tử có thể không đi qua toàn bộ chuỗi mà đi thẳng đến oxy, dẫn đến sự hình thành các superoxide mang gốc tự do. Đây là các phân tử hoạt tính hóa học mạnh có thể gây ra stress oxy hóa và góp phần vào lão hóa cũng như nhiều bệnh lý khác.
Các yếu tố oxy hóa khử trong ti thể
Năng lượng từ quá trình chuyển điện tử (mũi tên đen) trong chuỗi vận chuyển điện tử được sử dụng để bơm proton (mũi tên đỏ) từ chất nền của ti thể vào khoảng không giữa các lớp màng kép của ti thể. Điều này làm giảm tính axít và điện tích dương của chất nền ti thể, tạo ra sự chênh lệch pH và điện thế màng sinh chất giữa hai môi trường ngăn cách bởi màng ti thể, hình thành gradient proton điện hóa gọi là ΔΨ. Gradient điện hóa này tạo ra lực kéo proton có cường độ khoảng 180-190mV về phía chất nền. Proton di chuyển qua enzyme ATP synthase (ATP-ase), giúp tổng hợp ATP từ adenosine diphosphate (ADP) và phosphate vô cơ.
Các protein tham gia vào hệ thống vận chuyển điện tử chủ yếu được phân thành các phức hợp enzyme hô hấp chính. Phức hợp I (NADH coenzyme Q reductase) nhận điện tử từ NADH trong chu trình Krebs và chuyển đến coenzyme Q (ubiquinone, UQ). UQ cũng nhận điện tử từ phức hợp II (succinate dehydrogenase). UQ tiếp tục chuyển điện tử đến phức hợp III (cytochrome bc1), phức hợp III chuyển điện tử tới cytochrome c (cyt c). Cyt c đưa điện tử đến phức hợp IV (cytochrome c oxidase), nơi phức hợp IV sử dụng điện tử và proton H+ để khử oxy thành nước.
Có bốn phức hợp gắn trên màng sinh chất được xác định trong ti thể. Mỗi phức hợp là một cấu trúc phức tạp nằm trong lớp màng nội của ti thể. Ba trong số đó là các bơm proton. Các phức hợp này liên kết với nhau nhờ lực hút tĩnh điện giữa các chất mang điện tử hòa tan trong lipid. Tổng quan, chuỗi vận chuyển điện tử có thể được mô tả như sau:
NADH → Phức hợp I → Q → Phức hợp III → Cytochrome c → Phức hợp IV → O2 ↑ Phức hợp II
Phức hợp I
Tại Phức hợp I (NADH-CoQ Reductase, NADH dehydrogenase hay NADHoxidoreductase; EC 1.6.5.3), hai điện tử từ NADH được chuyển ra khỏi ion hydride. Vì phần lớn các chất mang chỉ có thể mang một điện tử tại một thời điểm, Flavin mononucleotide (FMN) trong phức hợp I hoạt động như một chất mang trung gian. FMN có khả năng chứa cả hai điện tử từ NADH nhưng chỉ nhận một điện tử mỗi lần và chuyển chúng vào nhân FeS của phức hợp I. Các điện tử tiếp tục được chuyển sang ubiquinone (Q), khử nó thành ubiquinol (QH2). QH2 di chuyển tự do trong màng sinh chất, trong khi phức hợp I bơm 4 proton (H) qua màng, tạo ra một gradient proton.
Phản ứng tổng quát do phức hợp I xúc tác là:
NADH + CoQ + 2H → NAD + H + CoQH2
Chu trình điện tử tại phức hợp I diễn ra như sau:
NADH được oxy hóa thành NAD qua việc khử Flavin mononucleotide thành FMNH2 trong một bước hai điện tử. FMNH2 sau đó bị oxy hóa qua hai bước một điện tử, với sự trung gian của semiquinone. Mỗi điện tử lần lượt được chuyển từ FMNH2 đến cụm sắt-lưu huỳnh và từ đó tới ubiquinone (Q). Sự chuyển giao điện tử đầu tiên tạo ra dạng gốc tự do semiquinone của Q, trong khi sự chuyển tiếp thứ hai khử semiquinone thành ubiquinol (QH2). Trong quá trình này, 4 proton H được bơm từ chất nền ti thể vào khoảng không giữa hai màng ti thể, tạo ra gradient proton giúp enzyme ATP synthease tổng hợp ATP qua phản ứng phosphorylate oxy hóa.
Phức hợp I là một trong bốn vị trí mà điện tử có thể rò rỉ tới ôxi, dẫn đến sự hình thành các superoxide độc hại.
Phức hợp II
Phức hợp II (Succinate - CoQ Reductase) chủ yếu bao gồm succinate dehydrogenase (EC 1..1). Enzyme này nhận các điện tử từ succinate và chuyển chúng vào quinone (Q) qua FAD. Tương tự như FMN, FAD có khả năng nhận một hoặc hai điện tử, nhưng chuyển từng điện tử một vào nhân FeS của phức hợp II, rồi FeS sẽ chuyển các điện tử này tới Q. Phản ứng tổng quát do phức hợp II xúc tác là:
Succinate + CoQ → Fumarate + CoQH2
Mặc dù phản ứng khử succinate thành fumarate có giá trị ΔG âm, năng lượng giải phóng không đủ để bơm proton từ chất nền ti thể vào khoảng không giữa hai lớp màng. Do đó, khác với các phức hợp I, III và IV, phức hợp II không bơm proton.
Phức hợp II cấu thành từ 4 đơn vị con: SDHA, SDHB, SDHC và SDHD. Một số nguồn cung cấp điện tử, như axít béo và glycerol 3-phosphate, cũng chuyển điện tử vào Q qua FAD.
Coenzyme Q
Coenzyme Q (ubiquinone) là một loại quinone với đuôi hydrocarbon dài, giúp phân tử CoQ dễ dàng khuếch tán trong lớp màng lipid của ti thể. Cả dạng khử (quinol) và dạng oxy hóa (quinone) của Q đều có khả năng di chuyển linh hoạt trong lớp màng lipid. Coenzyme Q nhận một điện tử từ phức hợp I hoặc II và di chuyển trong lớp màng ti thể cho đến khi gặp phức hợp III, nơi điện tử được chuyển tiếp. Sự di chuyển của CoQ và Cytochrome C trong lớp màng ti thể là yếu tố quan trọng giúp vận chuyển điện tử giữa các phức hợp trong chuỗi vận chuyển điện tử.
Phức hợp III
Tại Phức hợp III (Coenzyme Q - Cytochrome c Reductase, hay phức hợp cytochrome b-c1; EC 1.10.2.2), CoQH2 chuyển hai điện tử của nó vào phức hợp, trở lại dạng CoQ oxy hóa, đồng thời giải phóng hai proton vào khoảng không giữa hai lớp màng ti thể, tạo ra một phần của thế proton. Bên trong phức hợp, điện tử di chuyển từ CoQ đến nhân FeS, rồi tới một trong ba loại cytochrome (bL, bH, c1), và cuối cùng được chuyển sang cytochrome c, một phân tử ngoại biên hòa tan trong nước khuếch tán trong khoảng không giữa hai màng ti thể.
Phản ứng tổng quát mà phức hợp III xúc tác như sau:
2Cytochrome c3+ + CoQH2 → 2Cytochrome c + CoQ + 2H+
Mỗi cặp điện tử đi qua phức hợp III, phức hợp này bơm 2 proton vào khoảng không giữa hai màng ti thể.
Khi quá trình truyền điện tử bị biến đổi, như khi gặp một lớp màng sinh chất dày đặc hoặc một chất ức chế hô hấp như antimycin A, điện tử có thể bị rò rỉ tại phức hợp III và hình thành superoxide.
Cytochrome C
Cytochrome C là một protein lưu động quan trọng, hoạt động như một chất mang điện tử. Nó nằm trên bề mặt của màng trong ti thể, tiếp xúc với khoảng không giữa hai lớp màng và có nhiệm vụ truyền điện tử từ phức hợp III đến phức hợp IV. Sự di chuyển của Cytochrome C và CoQ trong màng ti thể rất quan trọng để đảm bảo điện tử được truyền đúng cách đến các phức hợp kế tiếp trong chuỗi vận chuyển điện tử.
Phức hợp IV
Tại Phức hợp IV (cytochrome c oxidase; EC 1.9.3.1), cytochrome c truyền điện tử của nó - mỗi lần một phân tử cytochrome c truyền một điện tử - đến phức hợp này. Điện tử đầu tiên đi vào một cặp ion đồng (Cua), sau đó chuyển sang cytochrome a, tiếp theo là một phức hợp chứa ion đồng Cub, rồi sang cytochrome a3, và cuối cùng đến phân tử oxy - vật nhận điện tử cuối cùng - để tạo thành nước.
Phản ứng tổng quát do phức hợp IV xúc tác là:
2Cyt c + 2H + 1/2O2 → 2Cyt c + H2O
Mỗi cặp điện tử được chuyển đến phức hợp IV dẫn đến việc phức hợp này bơm 2 proton vào khoảng không giữa hai lớp màng ti thể.
Bắt cặp với phosphorylate oxy hóa
Như đã nêu, chuỗi vận chuyển điện tử tạo ra sự chênh lệch thế điện hóa proton giữa hai môi trường bị ngăn cách bởi lớp màng trong ti thể. Điều này dẫn đến sự di chuyển của proton từ khoảng không giữa hai lớp màng ti thể trở lại chất nền để cân bằng sự chênh lệch đó. Dòng proton này đi qua một kênh ưa nước do ATP synthase tạo ra, kích hoạt quá trình tổng hợp ATP; năng lượng của proton được lưu trữ trong các phân tử ATP. Vì vậy, việc kết hợp giữa phosphorylate oxy hóa và sự chênh lệch thế điện hóa proton là điều kiện cần thiết để sản xuất ATP.
Trong một số tình huống, năng lượng từ chênh lệch thế proton không được sử dụng để tổng hợp ATP do sự hiện diện của các tác nhân tách cặp như 2,4-dinitrophenol (DNP). Những tác nhân này bắt proton ở khoảng không giữa hai lớp màng và trả proton trở lại chất nền ti thể mà không qua ATP synthase. Điều này dẫn đến việc lực vận động proton bị triệt tiêu và dù chuỗi vận chuyển điện tử vẫn hoạt động, không có ATP được tạo ra. Năng lượng từ thế proton được chuyển hóa hoàn toàn thành nhiệt năng, như trong các mô mỡ nâu, nơi giúp cung cấp nhiệt cho cơ thể. Sự điều tiết hô hấp cũng bị triệt tiêu bởi các protein này, khiến chuỗi vận chuyển điện tử hoạt động với tốc độ tối đa.
Sự điều tiết hoạt động của chuỗi vận chuyển điện tử
Hoạt động của chuỗi vận chuyển điện tử tại ti thể còn bị điều tiết bởi sự ức chế của thế điện hóa proton. Khi thế điện hóa cao xuyên qua lớp màng trong ti thể, chuỗi vận chuyển điện tử có thể bị ức chế hoặc di chuyển ngược lại. Các nghiên cứu cho thấy sự điều tiết này nhằm duy trì cân bằng giữa năng lượng tự do của việc bơm proton và việc chuyển điện tử; tức là độ lớn của thế proton ảnh hưởng đến tốc độ và hướng của chuỗi vận chuyển điện tử. Khi chênh lệch thế proton quá lớn, năng lượng cần để bơm proton ra khỏi chất nền cũng rất cao, làm đình trệ quá trình oxy hóa NAD, FADH2 và hoạt động của chuỗi vận chuyển điện tử.
Nồng độ ATP, ADP và phosphate vô cơ có ảnh hưởng gián tiếp đến hoạt động của chuỗi vận chuyển điện tử thông qua sự điều chỉnh hoạt động của ATP synthase. Khi tỷ lệ ATPgiảm do việc sử dụng ATP tăng cao, ATP synthase hoạt động mạnh mẽ hơn, cho phép nhiều proton đi qua và làm giảm thế điện hóa, kích thích tốc độ của chuỗi vận chuyển điện tử. Nếu nồng độ ADP quá thấp, thế điện hóa proton không thể được giảm qua quá trình tổng hợp ATP từ ADP, làm đình trệ chuỗi vận chuyển điện tử.
Như đã nêu, các protein tách cặp khiến thế proton và lực vận động proton bị tiêu hao nhanh chóng mà không cần đến ATP synthase. Sự sụp đổ của thế proton làm triệt tiêu các cơ chế điều tiết và chuỗi vận chuyển điện tử hoạt động hết công suất, không bị ảnh hưởng bởi nồng độ ADP hay ATP.
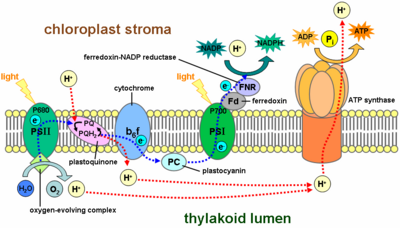
Chuỗi vận chuyển điện tử trong quá trình quang hợp
Trong quá trình phosphorylate oxy hóa, điện tử được truyền từ một chất cho điện tử có năng lượng cao (chẳng hạn như NADH) đến một chất nhận điện tử (ví dụ như O2) qua một chuỗi vận chuyển điện tử. Trong quá trình phosphorylate quang hóa, năng lượng ánh sáng mặt trời được sử dụng để tạo ra một chất cho điện tử có năng lượng cao và một chất nhận điện tử tương ứng. Điện tử được truyền từ chất cho đến chất nhận qua một chuỗi vận chuyển điện tử khác.
Chuỗi vận chuyển điện tử trong quang hợp có nhiều điểm tương đồng với chuỗi vận chuyển điện tử trong quá trình oxy hóa hô hấp. Chúng sử dụng các chất mang lưu động hòa tan trong lipid (như quinone) và các chất mang hòa tan trong nước (như cytochrome...). Chúng cũng bao gồm một bơm proton. Đặc biệt, bơm proton trong tất cả các chuỗi vận chuyển điện tử quang hợp đều tương tự như phức hợp số 3 trong ti thể.
Các chuỗi vận chuyển điện tử trong quang hợp sẽ được khám phá chi tiết hơn trong các bài về phosphorylate quang hóa, quang hợp, trung tâm phản ứng quang hợp và phản ứng phụ thuộc ánh sáng.
Kết quả
Chuỗi vận chuyển điện tử gồm các phản ứng oxy hóa khử, trong đó các electron được chuyển từ một tác nhân này sang một tác nhân khác. Quá trình chuyển electron làm thay đổi vị trí của proton qua màng sinh chất, tạo ra một gradient proton. Gradient này được sử dụng để cung cấp năng lượng cho các hoạt động cần thiết.
Sự kết hợp giữa các phản ứng sinh hóa thuận lợi và không thuận lợi (về mặt nhiệt động lực học) do các đại phân tử sinh học thực hiện là minh chứng cho tính chất mới phát sinh – một đặc tính không thể dự đoán trước, và ngay cả hiểu biết toàn diện về hệ địa hóa học cổ sơ từ nơi các đại phân tử đó tiến hóa cũng không thể giải thích. Câu hỏi mở là liệu những tính chất này xuất hiện ngẫu nhiên hay là kết quả tất yếu của quá trình tiến hóa trong các hệ thống sinh địa hóa học lớn, theo các định luật cơ bản của vật lý.
Chú thích
Ghi chú
Nguồn dẫn
- Giả thuyết CoRR
- Giả thuyết Hiđrô
- Bruce Alberts (2008). Phân Tử Sinh Học của Tế Bào. Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, Peter Walter. Garland Science, Taylor & Francis Group. ISBN 978-0-8153-4106-2.
- Harvey Lodish (2003). Sinh Học Tế Bào. Arnold Berk, Paul Matsudaira, Chris A. Kaiser, Monty Krieger, Matthew P. Scott, Lawrence Zipursky, James Darnell. ISBN 0716743663.
- H. P. Gajera, S. V. Patel, B. A. Golakiya (2008). Các Nguyên Lý Cơ Bản về Sinh Hóa: Sách Giáo Khoa. International Book Distributing Co. (Publishing Division). ISBN 978-81-8189-165-5.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- Fenchel T (2006). Hóa Sinh Vi Sinh: Sinh Thái Học của Chu trình Khoáng. King GM, Blackburn TH. Elsevier. ISBN 978-0121034559.
- Lengeler JW (1999). Sinh Học của Các Vi Khuẩn. Drews G; Schlegel HG (biên tập). Blackwell Science. ISBN 978-0632053575.
- Nelson DL (2005). Nguyên Lý Sinh Hóa Lehninger. Cox MM (ấn bản 4). W. H. Freeman. ISBN 978-0716743392.
- Nicholls DG (2002). Bioenergetics 3. Ferguson SJ. Academic Press. ISBN 978-0125181211.
- Stumm W (1996). Hóa Học Thủy Sinh. Morgan JJ (ấn bản 3). John Wiley & Sons. ISBN 978-0471511854.
- Thauer RK (1977). Jungermann K; Decker K. “Bảo Tồn Năng Lượng ở Vi Khuẩn Kị Khí Hóa Hóa”. Bacteriol Rev. 41 (1): 100–80. PMC 413997. PMID 860983.
- White D (1999). Sinh Lý và Sinh Hóa của Các Vi Khuẩn (ấn bản 2). Oxford University Press. ISBN 978-0195125795.
- Voet D (2004). Sinh Hóa Học. Voet JG (ấn bản 3). John Wiley & Sons. ISBN 978-0471586517.
- Kim HS.; Patel, K; Muldoon-Jacobs, K; Bisht, KS; Aykin-Burns, N; Pennington, JD; Van Der Meer, R; Nguyen, P; Savage, J (2010). “SIRT3 là một ức chế khối u nằm trong ty thể cần thiết để duy trì toàn vẹn và chuyển hóa ty thể trong điều kiện căng thẳng”. Cancer Cell. 17 (1): 41–52. doi:10.1016/j.ccr.2009.11.023. PMID 20129246.
Liên kết ngoài
- MeSH Chuỗi+Vận+Chuyển+Điện+Tử+Complex+Proteins
- UMich Hướng Dẫn Các Protein Trong Màng families/superfamily-3 - Các phức hợp với các miền cytochrome b-like
- UMich Hướng Dẫn Các Protein Trong Màng families/superfamily-4 - Cytochrome c oxidases ở vi khuẩn và ty thể
- UMich Hướng Dẫn Các Protein Trong Màng families/superfamily-2 - Các trung tâm phản ứng quang hợp và các hệ thống quang hợp
- UMich Hướng Dẫn Các Protein Trong Màng families/superfamily-78 - Gia đình Cytochrome c
- UMich Hướng Dẫn Các Protein Trong Màng families/superfamily-101 - Cupredoxins
- UMich Hướng Dẫn Các Protein Trong Màng protein/pdbid-1e6e - Adrenodoxin reductase
- UMich Hướng Dẫn Các Protein Trong Màng families/superfamily-130 - Flavoproteins truyền electron
Mẫu: Hô hấp tế bào Mẫu: Chuỗi vận chuyển electron
Cảnh báo: Từ khóa mặc định “Chuỗi_vận_chuyển_điện_tử” sẽ thay thế từ khóa trước đó, “Electron Transport Chain”.