

Trong sinh thái học, một khái niệm là sự phù hợp của một loài với một điều kiện môi trường cụ thể. Nó mô tả cách mà một sinh vật hoặc quần thể phản ứng với phân bố nguồn sống và các đối thủ cạnh tranh (ví dụ: bằng cách phát triển khi nguồn sống dồi dào và khi hiếm động vật ăn thịt, ký sinh và mầm bệnh) và cách mà nó thay đổi những yếu tố tương tự ấy (ví dụ: giới hạn tiếp cận nguồn sống bởi các sinh vật khác, đóng vai trò là nguồn thức ăn cho động vật ăn thịt và loài tiêu thụ con mồi). 'Loại và số lượng biến đổi là tập hợp gồm kích thước của một khu môi trường thay đổi từ loài này sang loài khác [và] tầm quan trọng tương đối của những biến đổi môi trường cụ thể với một loài có thể thay đổi tùy theo bối cảnh địa lý và sinh học'.
Khu Grinnel được xác định bởi sinh cảnh mà một loài sinh sống và những tập tính thích nghi đi kèm. Khu Elton nhấn mạnh rằng một loài không chỉ phát triển và phản ứng với môi trường, mà còn có thể thay đổi môi trường và tập tính khi nó lớn lên. Khu Hutchinson sử dụng toán học và thống kê để cố giải thích cách mà các loài cùng tồn tại trong một quần thể nhất định.
Khái niệm về khu sinh thái là trọng tâm của ngành địa sinh học sinh thái, chú trọng vào những mô hình không gian của các quần thể sinh thái. 'Sự phân bố của loài và động lực học của chúng theo thời gian là kết quả của những đặc tính của loài, biến đổi môi trường..., và tương loài giữa hai loài—cụ thể là khả năng của một số loài (đặc biệt là loài người chúng ta), nhằm điều chỉnh môi trường của chúng và làm thay đổi động lực phạm vi của nhiều loài khác.' Sự thay đổi của một khu sinh thái bởi sinh vật cư ngụ là đề tài của thiết kế khu.
Phần lớn các loài tồn tại trong một môi trường sinh thái chuẩn, chia sẻ các tập tính, thích nghi và chức năng tương tự như những loài có quan hệ chặt chẽ khác ở cùng một hạng, mặc dù vẫn có một số ngoại lệ. Một ví dụ điển hình về loài không tuần hoàn là chim kiwi ở New Zealand; chim này ăn giun và các sinh vật khác trên mặt đất, đồng thời sinh sống trong một tổ như động vật có vú. Địa sinh học đảo có thể giải thích các loài trên đảo và những tổ không tuần hoàn.
Grinnell Nest
Ý nghĩa sinh thái của ổ bắt nguồn từ khái niệm ổ là một cái lỗ để đặt bức tượng, có nguồn gốc từ từ nicher tiếng Pháp cổ, nghĩa là nest. Thuật ngữ này được nhà tự nhiên học Roswell Hill Johnson đề xuất, nhưng Joseph Grinnell là người đầu tiên sử dụng trong một nghiên cứu năm 1917, trong bài báo 'Mối quan hệ ổ của chim họa mi đỏ California' của ông.
Khái niệm ổ Grinnell giới thiệu ý tưởng rằng một loài được định nghĩa bởi môi trường sống và các tập tính thích nghi đi kèm. Nói cách khác, ổ là tổng hợp yêu cầu sinh cảnh và tập tính cho phép một loài tồn tại và sinh sản. Ví dụ, chim họa mi đỏ California phù hợp với môi sinh chaparral mà nó sinh sống—sinh sản và kiếm ăn trong bụi rậm và tránh lũ quét bằng cách di chuyển từ bụi này sang bụi khác. 'Ổ' của chim được xác định bằng sự kết hợp hợp lý giữa tập tính và đặc tính hình thái của chim (màu lông, cánh ngắn, chân mạnh) với môi sinh này.

Ổ Grinnell có thể được xác định bởi sự thay đổi phi tương tác (phi sinh học) và các điều kiện môi trường trên quy mô lớn. Các thay đổi quan trọng trong loại ổ này bao gồm nhiệt độ trung bình, lượng mưa, bức xạ mặt trời và các khía cạnh của địa hình, được ngày càng dễ tiếp cận trên quy mô không gian. Phần lớn tài liệu chú trọng vào cấu trúc ổ Ginnell, thường từ góc độ khí hậu, để giải thích sự phân bố và sự phong phú. Các dự đoán hiện tại về phản ứng của loài với biến đổi khí hậu chủ yếu dựa trên những dự đoán về sự thay đổi môi trường đối với sự phân bố của loài. Tuy nhiên, giới khoa học ngày càng nhận thức rõ hơn về việc biến đổi khí hậu cũng ảnh hưởng đến tương tác giữa các loài và góc nhìn của Elton có thể dễ dàng giải thích những quá trình này.
Góc nhìn này của ổ cho phép tồn tại cả các đơn vị sinh thái tương đương và những ổ trống. Một đơn vị sinh thái tương đương với một sinh vật là một sinh vật từ một nhóm phân loại khác thể hiện sự thích nghi tương tự trong cùng một môi trường sống, ví dụ như thực vật khác nhau của sa mạc, xương rồng và đại kích ở Mỹ và châu Phi. Một ví dụ khác, thằn lằn anole ở Đại Antilles là một ví dụ hiếm hoi về tiến hóa hội tụ, sự thích ứng bức xạ và sự tồn tại của đơn vị sinh thái tương đương: thằn lằn anole tiến hóa trong cùng một môi trường sống độc lập với nhau và tạo ra hình thái sinh thái giống nhau trên cả bốn hòn đảo.
Ổ Elton
Năm 1927, nhà sinh thái học người Anh Charles Sutherland Elton đã định nghĩa ổ như sau: ''Ổ' của một động vật là vị trí của nó trong môi trường sống, mối quan hệ của nó với thức ăn và kẻ thù.'
Elton phân loại ổ theo các hoạt động săn mồi ('tập tính săn mồi'):
Ví dụ, có một ổ bị lấp đầy bởi các loài chim săn mồi nhỏ như chuột chù và chuột nhắt. Trong một khu rừng sồi, ổ này bị chiếm đóng bởi cú vàng nâu, trong khi ở đồng cỏ trống, nó lại thuộc về chim cắt. Sự tồn tại của ổ sinh vật ăn thịt này phụ thuộc vào thực tế rằng chuột nhắt hình thành một ổ động vật ăn cỏ rõ ràng trong nhiều mối liên hệ khác nhau, bất kể loài chuột nhắt có thể khác nhau.

Về mặt khái niệm, ổ Elton giới thiệu ý tưởng về phản ứng và tác động của một loài đối với môi trường. Khác với các khái niệm ổ khác, nó nhấn mạnh rằng một loài không chỉ phát triển và phản ứng với môi trường dựa trên nguồn sống, động vật ăn thịt và điều kiện khí hậu có sẵn, mà còn điều chỉnh sự hiện diện và hành vi của những yếu tố đó khi nó phát triển. Ví dụ điển hình, hải ly cần một số nguồn sống cụ thể để tồn tại và sinh sản, nhưng việc xây dựng các đập cũng làm thay đổi dòng chảy của dòng sông nơi chúng sống. Do đó, hải ly ảnh hưởng đến điều kiện sinh học và phi sinh học của các loài khác sống trong và gần với chúng. Trong một trường hợp ít gặp hơn, các sinh vật cạnh tranh tiêu thụ các nguồn sống ở tỷ lệ khác nhau có thể dẫn đến chu kỳ biến động mật độ nguồn sống giữa các loài. Các loài không chỉ phát triển khác nhau liên quan đến mật độ nguồn sống, mà sự gia tăng quần thể của chính chúng có thể ảnh hưởng đến mật độ nguồn sống theo thời gian.
Ổ Elton tập trung vào các tương tác sinh học và động lực sinh vật ăn thịt-nguồn sống (các thay đổi sinh học) trên quy mô địa phương. Vì phạm vi tập trung hẹp, các bộ dữ liệu đặc trưng cho ổ Elton thường là nghiên cứu thực địa chi tiết về các hiện tượng cụ thể, vì tính động lực của lớp ổ này khó đo lường ở quy mô địa lý rộng. Tuy nhiên, ổ Elton có thể hữu ích trong việc giải thích khả năng chịu đựng thay đổi toàn cầu của một loài. Do những điều chỉnh trong các tương tác sinh học sẽ dẫn đến thay đổi các yếu tố phi sinh học, vì vậy ổ Elton có thể hữu ích trong việc miêu tả phản ứng tổng thể của một loài đối với môi trường mới.
Ổ Hutchinson

Ổ Hutchinson là một 'siêu không gian chiều n' trong đó các chiều đại diện cho điều kiện môi trường và nguồn sống, quyết định yêu cầu để tồn tại của cá thể hoặc loài, mà cụ thể hơn là sự tồn tại của cả quần thể. Khái niệm 'siêu không gian' xác định không gian đa chiều của nguồn sống (như ánh sáng, chất dinh dưỡng, cấu trúc,...) và được các sinh vật sử dụng, và 'mọi loài khác đều được coi là một phần của hệ thống tọa độ.'
Khái niệm ổ sinh vật được G. Evelyn Hutchinson đưa ra vào năm 1957. Hutchinson quan tâm đến lý do tại sao có quá nhiều loài sinh vật tồn tại trong bất kỳ môi trường nào. Công trình của ông đã truyền cảm hứng cho nhiều sinh vật học khác phát triển các mô hình để giải thích số lượng và sự đa dạng của các loài cùng tồn tại trong một quần thể nhất định và dẫn đến các khái niệm về 'rộng rãi ổ' (sự đa dạng của các nguồn sống hoặc môi trường sinh sống mà một loài sử dụng), 'phân hóa ổ' (phân hoá nguồn sống theo các loài cùng tồn tại) và 'chồng chéo ổ' (sự chồng chéo giữa các nguồn sống mà các loài khác nhau sử dụng).
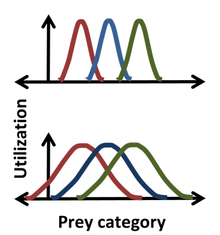
Robert MacArthur và Richard Levins đã sử dụng các số liệu thống kê vào ổ Hutchinson bằng cách sử dụng ổ 'nguồn sống-tận dụng' để mô tả 'tần suất xảy ra' như là một hàm của tọa độ Hutchinson. Ví dụ, một phân phối Gauss có thể mô tả tần suất mà một loài săn mồi có kích thước nhất định, cho phép mô tả chi tiết hơn về ổ hơn là chỉ ghi lại một vài số trung vị hoặc kích thước trung bình của con mồi. Đối với phân bố hình chuông như vậy, vị trí, phạm vi và hình dạng của ổ tương ứng với giá trị trung bình, độ lệch chuẩn và chính phương pháp phân phối thực tế của ổ. Một lợi thế khi sử dụng số liệu thống kê được minh họa trong hình vẽ, nơi mà rõ ràng rằng trong các khu vực phân bố hẹp hơn (phía trên), không có sự cạnh tranh giữa các loài săn mồi từ hai bên trái và phải, trong khi đối với các khu vực phân bố rộng hơn (phía dưới), sự chồng chéo của ổ chỉ ra rằng sự cạnh tranh có thể xảy ra giữa tất cả các loài. Cách tiếp cận nguồn sống-tận dụng cho rằng sự cạnh tranh không chỉ có thể xảy ra mà còn làm xảy ra, và sự chồng chéo trong việc sử dụng nguồn sống giữa các loài cho phép ước tính các hệ số cạnh tranh. Tuy nhiên, nguyên lý này có thể dẫn đến nhầm lẫn khi bỏ qua các tác động của nguồn sống của mỗi loài lên sinh vật và tác động của sinh vật lên nguồn sống của từng loài. Ví dụ, trong các vùng phân bố chồng chéo, nguồn sống có thể không bị giới hạn, do đó không có sự cạnh tranh cho nguồn sống này dù có sự chồng chéo ổ.

Một sinh vật không có sự can thiệp từ các loài khác có thể sử dụng đầy đủ điều kiện (sinh học và phi sinh học) và các nguồn sống để tồn tại và sinh sản được gọi là ổ cơ bản. Tuy nhiên, do áp lực và tương tác với các loài khác (cạnh tranh giữa các loài), sinh vật thường phải chiếm giữ một không gian hẹp hơn và có sự thích nghi cao; đây là ổ xác định rộng. Hutchinson đã sử dụng khái niệm cạnh tranh nguồn sống như một cơ chế chính điều khiển hệ sinh thái, nhưng việc tập trung quá nhiều vào điều này đã trở thành một trở ngại đối với khái niệm ổ. Đặc biệt là việc nhấn mạnh quá mức vào sự phụ thuộc của một loài vào nguồn sống có thể dẫn đến việc ít chú ý đến tác động của sinh vật lên môi trường của chúng, ví dụ như chiếm giữ và xâm lấn.
Thuật ngữ 'vùng thích nghi' được đưa ra bởi nhà cổ sinh vật học George Gaylord Simpson để giải thích cách mà một quần thể có thể di chuyển từ một ổ này sang ổ khác phù hợp với nó, chuyển sang 'vùng thích ứng', có thể được điều chỉnh hoặc thay đổi trong chuỗi thức ăn, làm cho vùng thích nghi phù hợp với nó mà không gặp sự gián đoạn trong cách sống của nó do nhóm đã 'thích nghi trước' với cơ hội sinh thái mới.
Một ổ là một phần rất đặc biệt của không gian sinh thái được một loài độc chiếm. Với giả định rằng không có hai loài giống nhau mặt mũi (được gọi là 'tiên đề bất bình đẳng' của Hardin) và nguyên lý loại trừ cạnh tranh, một số khía cạnh của nguồn sống hoặc thích nghi sẽ cung cấp một ổ cụ thể cho từng loài. Tuy nhiên, các loài có thể chung 'lối sống' hoặc 'chiến lược sinh thái đơn loài', định nghĩa mở rộng hơn về không gian sinh thái. Ví dụ, những loài ở đồng cỏ Úc (mặc dù khác biệt so với các loài ở đồng cỏ Bắc Mỹ) vẫn thể hiện những lối sống tương tự nhau.
Biểu diễn toán học về ổ cơ bản của một loài trong không gian sinh thái và phép chiếu của nó trở lại không gian địa lý là lĩnh vực của mô hình ổ.
Thuyết hiện đại về mối quan hệ sinh vật
Thuyết hiện đại về mối quan hệ sinh vật (hay còn gọi là 'thuyết hiện đại về mối quan hệ sinh thái' trong một số ngữ cảnh) là một khung lý thuyết ban đầu được thiết kế để hòa hợp các định nghĩa khác nhau về môi trường sống (xem các định nghĩa của Grinnell, Elton và Hutchinson ở trên) và giúp giải thích các quá trình cơ bản tác động đến hệ sinh thái theo mô hình Lotka–Volterra. Khung lý thuyết tập trung vào 'mô hình nguồn sống-sinh vật tiêu thụ', chia hệ sinh thái thành các nguồn sống nhất định (ví dụ: ánh sáng mặt trời hoặc nước trong đất) và sinh vật tiêu thụ (bao gồm cả thực vật và động vật) và cố gắng xác định phạm vi các mối quan hệ có thể tồn tại giữa hai nhóm.
Trong thuyết hiện đại về mối quan hệ sinh vật, 'mối quan hệ tác động' được định nghĩa là sự kết hợp những tác động mà một sinh vật tiêu thụ cụ thể gây ra đối với cả nguồn sống mà nó sử dụng lẫn sinh vật tiêu thụ khác trong hệ sinh thái. Do đó, mối quan hệ tác động tương đương với mối quan hệ Elton vì cả hai khái niệm đều dựa trên tác động của một loài nhất định lên môi trường của nó.
Phạm vi các điều kiện môi trường nơi một loài có thể tồn tại và sinh sản thành công (tức là định nghĩa của Hutchinson về một môi trường sống cụ thể) cũng được tích hợp vào thuyết hiện đại về mối quan hệ sinh vật, gọi là 'môi trường sống yêu cầu'. Môi trường sống yêu cầu bị giới hạn bởi sự hiện diện của nguồn sống và tác động của sinh vật tiêu thụ cùng tồn tại (ví dụ: các loài cạnh tranh và loài săn mồi).
Tương đồng với thuyết hiện đại về mối quan hệ sinh vật
Thuyết về sự cùng tồn tại của hai loài sinh vật trong một môi trường nhất định đặt ra ba điều kiện quan trọng:
- Cả hai loài phải chia sẻ các yêu cầu sống chồng chéo.
- Mỗi loài phải cạnh tranh để giành nguồn sống cần thiết nhất. Ví dụ, nếu hai cây (P1 và P2) đấu tranh cho nitơ và phosphor trong một hệ sinh thái, họ chỉ có thể cùng tồn tại nếu chúng bị hạn chế bởi nguồn sống khác nhau (P1 có thể bị hạn chế bởi nitơ trong khi P2 bị hạn chế bởi phosphor) và mỗi loài phải chiếm ưu thế để thu được nguồn sống đó (P1 cần nitơ hơn P2 cần phosphor hơn). Điều này ngược lại có nghĩa là nếu cả hai đều cạnh tranh cùng một nguồn sống, loài chiến thắng sẽ sống sót và loài kia sẽ mất. Ngoài ra, nếu P1 bị cạnh tranh về nitơ (nguồn sống cần thiết nhất), nó sẽ không thể tồn tại. Tương tự, nếu P2 bị cạnh tranh về phosphor, nó cũng sẽ không thể sống sót.
- Sự hiện diện của các nguồn sống hạn chế (nitơ và phosphor trong ví dụ trên) trong môi trường phải tương đương nhau.
Những yêu cầu này đều rất thú vị và gây tranh cãi vì chúng yêu cầu cả hai loài phải có chung một môi trường nhất định (với các yêu cầu sống chồng chéo), nhưng cách chúng tương tác với môi trường đó lại khác nhau. Những yêu cầu này thường bị vi phạm khi các loài không bản địa (những loài xâm nhập) sống cùng với các loài địa phương mới trong phạm vi không phải là môi trường bản địa của chúng, nhưng có vẻ như các yêu cầu này không bị hạn chế. Nói cách khác, thuyết về sự cùng tồn tại đương đại dự đoán rằng các loài không thể xâm chiếm các môi trường mới ngoài vùng yêu cầu sống (hay vùng xác định), nhưng đã có nhiều ví dụ để minh chứng cho sự vi phạm của giả thuyết này. Ngoài ra, thuyết đương đại dự đoán rằng các loài không thể tồn tại trong môi trường mà các loài khác đã cạnh tranh nguồn sống giống như các loài sắp đến, nhưng cũng có nhiều ví dụ để hỗ trợ giả thuyết này.
Phạm vi địa lý và vùng sống
Phạm vi địa lý của một loài có thể coi là không gian mà nó chiếm giữ, cùng với các đặc điểm của mô hình địa lý và loài ảnh hưởng đến khả năng định cư của nó. Phạm vi địa lý cơ bản của một loài là vùng mà nó chiếm giữ trong điều kiện môi trường thuận lợi, không bị hạn chế từ các yếu tố phân tán hoặc định cư. Một loài sẽ bị giới hạn trong phạm vi địa lý xác định của nó khi phải đối mặt với tương tác sinh học hoặc rào cản phi sinh học hạn chế phân tán, là một bộ phận hẹp hơn của phạm vi địa lý cơ bản của nó.
Một nghiên cứu ban đầu về sinh thái ổ bởi Joseph H. Connell đã phân tích các yếu tố môi trường làm hạn chế phạm vi sống của loài hàu biển (Chthamalus stellatus) tại Đảo Cumbrae, Scotland. Trong nghiên cứu của mình, Connell đã mô tả các đặc điểm nổi bật của ổ C. stellatus và cung cấp giải thích về phân bố của chúng trên bãi triều đá của bờ biển. Connell chỉ ra rằng phạm vi phân bố của C. stellatus bị giới hạn bởi khả năng chịu mất nước trong khoảng thời gian triều thấp. Phần dưới của phạm vi bị giới hạn bởi các tương tác giữa các loài, đặc biệt là sự cạnh tranh với một loài hàu sống cùng và sự săn mồi của một con ốc sên. Bằng cách loại bỏ sự cạnh tranh từ B. balanoides, Connell đã chứng minh rằng C. stellatus có thể mở rộng phạm vi dưới trong trường hợp không có sự loại trừ cạnh tranh. Những thí nghiệm này minh chứng cho vai trò của các yếu tố sinh học và phi sinh học trong việc hạn chế phân bố của một sinh vật.
Các tham số
Các kích thước khác nhau hoặc các trục cắt của biểu đồ, đại diện cho các biến số sinh học và phi sinh học khác nhau. Những yếu tố này có thể bao gồm miêu tả về lịch sử sống, sinh cảnh, vị trí dinh dưỡng (trong chuỗi thức ăn) và phạm vi địa lý của sinh vật. Theo nguyên lý loại trừ cạnh tranh, không có hai loài nào có thể chia sẻ cùng một ổ trong cùng một môi trường trong thời gian dài. Các tham số của một ổ xác định được mô tả bằng sự rộng của nó. Một số loài thực vật và động vật (được gọi là loài chuyên hóa) cần môi trường và sinh cảnh cụ thể để tồn tại, ví dụ như cú lông đốm, loài chuyên sống trong rừng già. Các loài thực vật và động vật khác (được gọi là loài rộng sinh thái) không chuyên hóa và có thể tồn tại trong nhiều điều kiện, như bồ công anh.