Trong lĩnh vực sinh học phân tử, quá trình nhân đôi DNA hay tổng hợp DNA là quá trình sao chép các phân tử DNA xoắn kép trước mỗi lần phân bào. Kết quả của quá trình này là tạo ra hai phân tử DNA gần như giống nhau hoàn toàn, chỉ sai khác với tần số rất thấp (thông thường dưới một phần vạn, xem thêm đột biến). Cơ chế nhân đôi thực hiện dựa trên nguyên tắc bổ sung, và tế bào có hệ thống tìm kiếm và sửa chữa các sai sót DNA một cách hiệu quả, có chuyên môn, tỉ mỉ, cẩn trọng nhưng vẫn chưa thể hoàn toàn tránh khỏi các đột biến. Hiện nay, các cấu trúc của phân tử RNA có thể dễ dàng bị phá vỡ, ảnh hưởng đến cấu trúc DNA khác một cách nghiêm trọng, gây hại đến sức khỏe của chúng ta.
Nguyên lí cơ bản
Quá trình nhân đôi DNA ở tế bào đơn bào, đa bào và DNA của virut (dạng sợi kép) đều theo nguyên tắc bổ sung và bảo toàn.
Chứng minh quá trình nhân đôi DNA diễn ra theo nguyên tắc bảo toàn: phương pháp sử dụng đồng vị phóng xạ.
Quá trình nhân bản DNA
Phần mở đầu
Để một tế bào có thể chia đôi, trước tiên nó phải nhân đôi DNA của mình. [11] Quá trình này bắt đầu tại các điểm cụ thể trên DNA, được đích thị bởi các protein khởi đầu. [4] Trong E. coli, protein này là DnaA; trong nấm men, nó là một phức hợp nhận diện gốc. [12] Chuỗi DNA mục tiêu của protein khởi đầu có xu hướng 'giàu A, T' (adenine và thymine), vì các cặp A-T có hai liên kết hydro (thay vì ba như trong cặp G-X) và dễ dàng tách rời hơn. [13] Khi điểm bắt đầu đã được xác định, những protein này tạo thành phức hợp tiền mẫu và làm nở hai sợi DNA.
Phần kéo dài
Polymeraza DNA có hoạt động theo hướng 5′ –3. Tất cả các hệ thống sao chép DNA đều có một nhóm hydroxyl tự do ở 3' của mạch trước khi tổng hợp có thể bắt đầu (lưu ý: mẫu DNA được đọc theo hướng 3′ đến 5' trong khi chuỗi mới được tổng hợp theo hướng 5′ đến 3' — thường là đoạn lên của chuỗi). Bốn cơ chế riêng biệt cho tổng hợp DNA đã được xác nhận:
- Tất cả các loại tế bào sống và nhiều loại virus DNA, các phage và plasmid đều sử dụng một enzym primaza để tổng hợp một mồi RNA ngắn có một nhóm 3′ OH tự do, sau đó được kéo dài bởi DNA polymeraza.
- Các retroelement (bao gồm cả retrovirus) sử dụng một RNA chuyển đổi để sao chép DNA bằng cách cung cấp một 3 ′OH tự do được sử dụng để kéo dài bởi enzym phiên mã ngược.
- Trong Virus Adeno và họ φ29 của vi khuẩn, nhóm 3 'OH được cung cấp bởi chuỗi bên của một acid amin bộ gen gắn protein (protein đầu cuối) mà nucleotide được DNA polymeraza thêm vào để tạo thành một chuỗi mới.
- Trong các virus DNA mạch đơn - một nhóm bao gồm các circovirus, geminivirus, parvovirus và các loại khác - cũng như nhiều loại thực thể và plasmid sử dụng cơ chế nhân rộng vòng tròn lăn (RCR), endonuclease RCR tạo ra một vết cắt trong chuỗi gen (virus đơn mạch) hoặc một trong hai mạch DNA (plasmid). Đầu 5' của mạch có vết này được chuyển sang một tyrosine dư thừa trên nucleaza và nhóm 3 ′OH tự do sau đó được DNA polymeraza sử dụng để tổng hợp chuỗi mới.
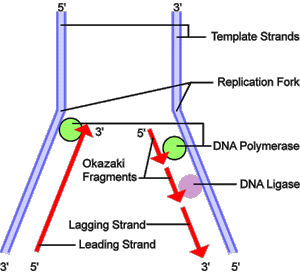
Đầu tiên, cơ chế này được biết đến nhiều nhất và được sử dụng bởi các sinh vật di động. Trong cơ chế này, sau khi hai mạch được tách ra, primaza bổ sung một mồi RNA vào mỗi mạch mẫu. Mạch dẫn đầu nhận một mồi RNA trong khi mạch trễ nhận một số mồi. Mạch dẫn đầu được kéo dài liên tục từ mồi bằng một DNA polymeraza với xử lý nhanh, trong khi mạch trễ được kéo dài không liên tục thành các đoạn Okazaki. RNase loại bỏ các đoạn mồi RNA và một DNA polymeraza khác với xử lý chậm được sử dụng để điền vào khoảng trống. Khi hoàn thành, một vết đơn lẻ có thể được tìm thấy trên mạch mẫu và một số vết trên mạch kia. Ligaza tham gia để lấp đầy các vết này, hoàn thành quá trình nhân đôi của phân tử DNA mới.
Primaza sử dụng trong quá trình này có sự khác biệt đáng kể giữa vi khuẩn và vi sinh vật / sinh vật nhân chuẩn. Vi khuẩn sử dụng primaza thuộc họ siêu protein DnaG chứa một tên xúc tác của loại gấp TOPRIM. [14] Vòng TOPRIM chứa một lõi α / β với bốn sợi được bảo tồn trong một cấu trúc gắn kết tương tự như Rossmann. Cấu trúc này cũng được tìm thấy trong các lĩnh vực xúc tác của topoisomerase Ia, topoisomerase II, nucleaza và protein sửa chữa DNA của họ OLD liên quan đến protein RecR.
Các enzym tham gia
Gyraza (hay còn gọi là topoisomeraza II): Làm thẳng phân tử DNA.
Helicaza: giải cấu trúc xoắn của DNA và giải thể hai mạch đơn bằng cách cắt các liên kết hydro.
DNA polymeraza:
DNA polymeraza I: loại bỏ mạch RNA khởi đầu và tổng hợp mạch polinucleotide mới.
DNA polymeraza II: sửa chữa sai sót sau khi nối các mảnh Okazaki.
DNA polymeraza III: lắp ráp nòng nuôi và kéo dài mạch đơn mới.
Ligaza: kết nối các mảnh Okazaki.
Primaza (RNA polymeraza): Tổng hợp mảnh mồi.
Ngoài ra còn có:
Prôtêin SSB: giúp hai mạch đơn không bị dính lại với nhau để các enzym có thể hoạt động.
Telomeraza: hạn chế sự cố đầu mút. Chỉ tồn tại trong tinh hoàn và buồng trứng, không hoạt động trong tế bào sinh dưỡng.
Diễn tiến
Trong tế bào sinh vật đơn bào
Bước 1: Giải mã xoắn phân tử DNA
Nhờ enzym tôpôizômêraza giải mã xoắn, phân tử DNA khỏi cấu trúc siêu xoắn của tôpô, sau đó enzym hêlicaza giải hai mạch đơn của DNA, tạo thành hình chữ Y để lộ ra hai đoạn mạch đơn làm khuôn.
Bước 2: Tổng hợp mạch DNA mới
Enzyme DNA polymerase sử dụng 2 mạch đơn mẫu để tổng hợp mạch mới theo nguyên tắc bổ sung.
Do enzyme DNA polymerase chỉ có thể tổng hợp mạch mới theo chiều 5' - 3' (chỉ khi có đầu 3' OH tự do), vì vậy trên mạch mẫu có chiều 3'-5', quá trình tổng hợp mạch mới diễn ra một cách liên tục. Mạch này được gọi là mạch dẫn đầu.
Sau đó, các đoạn Okazaki được nối lại với nhau bằng enzyme nối ligaza.
Bước 3: Hai phân tử DNA hoàn thành (kết thúc)
Mạch mới được tổng hợp theo một mạch đơn, trong khi mạch còn lại đến từ DNA ban đầu (nguyên tắc bảo toàn).
Tuy nhiên, các tế bào sinh vật nhân thực thường có nhiều phân tử DNA có kích thước lớn. Quá trình nhân đôi DNA xảy ra ở nhiều vị trí trên mỗi phân tử DNA tạo ra nhiều đơn vị tái bản với sự tham gia của nhiều loại enzym khác nhau.
Ở sinh vật nhân thực, quá trình tổng hợp mạch mới bắt đầu từ đầu mạt của phân tử DNA thường có một hiện tượng đặc biệt được gọi là sự cố đầu mạt.
Kết quả của quá trình nhân đôi ADN
Kết quả của quá trình nhân đôi là tạo ra 2 phân tử ADN con từ một phân tử ADN mẹ.
Phân tử ADN con có những đặc điểm sau:
- Thường thì, hai phân tử ADN con có cấu trúc gần giống hệt cấu trúc của phân tử ADN mẹ, trong một số trường hợp có những khác biệt rất nhỏ, không đáng kể.
- Trên phân tử ADN con cũng có 2 chuỗi xoắn nhưng chỉ có một mạch đơn được tổng hợp mới, mạch còn lại là mạch đơn từ ADN mẹ.
Hình ảnh