

| Dực long | |
|---|---|
| Khoảng thời gian tồn tại: Tam Điệp muộn – Phấn Trắng muộn, 228–66 triệu năm trước đây TiềnЄ
Є
O
S
D
C
P
T
J
K
Pg
N
| |
| Bản phục dựng khung xương của loài Geosternbergia sternbergi, con cái (trái) và con đực (phải) | |
| Phân loại khoa học | |
| Vực: | Eukaryota |
| Giới: | Animalia |
| Ngành: | Chordata |
| Lớp: | Reptilia |
| nhánh: | Ornithodira |
| nhánh: | †Pterosauromorpha |
| Bộ: | †Pterosauria Kaup, 1834 |
| Các phân nhóm | |
| |
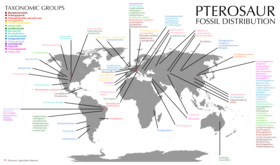
| Phân bố hóa thạch dực long. Các loài và danh pháp được tô màu tương ứng với phân loại của chúng. | |
Thằn lằn có cánh, còn được gọi là thằn lằn bay hay dực long, là một nhóm bò sát biết bay thuộc nhánh/bộ Pterosauria. Chúng tồn tại từ Kỷ Tam Điệp muộn đến cuối kỷ Phấn Trắng (từ 210 đến 66 triệu năm trước). Pterosauria là động vật có xương sống đầu tiên thích ứng cho việc bay. Cánh của chúng được cấu tạo từ một màng da (giống như cánh dơi), cơ và các tế bào khác kéo dài từ chân đến ngón thứ tư kéo dài. Các loài cổ nhất có răng hàm dài, trong khi các loài xuất hiện sau có đuôi bị thoái hóa, và một số loài không có răng. Một số loài có lớp lông giống như tóc, che phủ thân và một phần cánh của chúng. Dực long có kích thước đa dạng từ rất nhỏ (như Nemicolopterus) đến rất lớn (như Quetzalcoatlus và Hatzegopteryx).
Pterosauria đôi khi cũng được gọi là pterodactyl, đặc biệt là trên các phương tiện truyền thông. Tuy nhiên, thuật ngữ này không được khuyến khích sử dụng. 'Pterodactyl' ám chỉ các loài trong chi Pterodactylus và rộng hơn là các loài trong phân bộ Pterodactyloidea.
Mô tả
Giải phẫu của loài thằn lằn bay cho thấy sự biến đổi đáng kể so với tổ tiên bò sát của chúng để phù hợp với việc bay. Xương dực long giống xương chim, rỗng và chứa không khí. Chúng có xương ức cong lưỡi hái phát triển để các cơ bắp hỗ trợ bay có nhiều diện tích bám vào hơn và một bộ não lớn chuyên hóa cho việc bay. Ở một số loài dực long sau này, xương vai hợp nhất với xương sống thành một cấu trúc duy nhất gọi là xương notarium, làm cứng thân trong chuyến bay và gia cố ổn định cho hai xương vai.
Đôi cánh
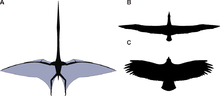
Cánh của thằn lằn bay cấu tạo từ màng da và các loại mô khác. Các màng chính gắn vào ngón tay thứ tư cực dài của mỗi cánh tay và kéo dài dọc theo hai bên cơ thể đến mắt cá chân.
Ban đầu, cánh chỉ được coi là một cấu trúc da đặc biệt cho đến khi các nghiên cứu sau này chỉ ra rằng màng cánh của thằn lằn bay là một cấu trúc cơ sinh học rất phức tạp, phù hợp cho lối sống bay lượn. Phần cánh ngoài (từ đầu ngón tay đến khuỷu tay) được tăng cường bởi các dây chằng so le gọi là actinofibril. Các sợi actinofibril tạo thành ba lớp riêng biệt ở cánh, tạo nên một mô hình lồng ghép. Chức năng và vật liệu của các sợi actinofibril chưa được xác định rõ, nhưng có thể giúp gia cố hoặc bổ sung năng lượng cho cánh. Màng cánh cũng chứa một lớp cơ mỏng, mô xơ và một hệ tuần hoàn với các mạch máu vòng lặp phức tạp, độc đáo.
Xương cánh của dực long có lỗ hổng khá lớn và mô mềm được bảo quản trong ít nhất một mẫu. Một số loài thằn lằn bay có hệ thống các túi khí hô hấp mở rộng (xem phần Cổ sinh thái học bên dưới) để tạo thành màng cánh.
Các phần của đôi cánh

Màng cánh của thằn lằn bay được chia thành ba phần chính. Thứ nhất, màng trước (propatagium) là phần đầu của cánh, gắn từ cổ tay đến vai, tạo ra 'cạnh dẫn đầu' khi bay. Màng này có thể đã kết nối ba ngón đầu tiên của bàn tay, được minh chứng trong một số mẫu vật. Thứ hai, 'màng cánh tay' (brachiopatagium) là thành phần chính, kéo dài từ ngón tay thứ tư đến chân sau (mặc dù vị trí chính xác vẫn đang tranh cãi và có thể khác nhau giữa các loài). Cuối cùng, một số loài có màng kéo dài giữa hai chân, có thể kết nối với đuôi, gọi là uropatagium; mức độ trải dài chưa được xác định rõ, nghiên cứu về chi Sordes cho thấy nó chỉ kết nối với chân, không liên quan đến đuôi (tạo thành cruropatagium). Các nhà khoa học đồng ý với nghiên cứu này, mặc dù các loài Rhamphorhynchoidea có uro/cruropatagium rộng hơn, còn pterodactyloid chỉ có màng chạy dọc chân.
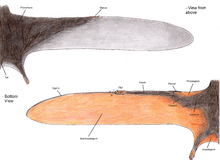
Xương pteroid chỉ có ở thằn lằn bay, kết nối với cổ tay và hỗ trợ màng propatagium giữa cổ tay và vai. Bằng chứng về màng kết nối ba ngón tay trước của thằn lằn bay cho thấy màng này có thể đã mở rộng hơn so với kết nối pteroid-vai thường thấy. Vị trí của xương pteroid vẫn còn gây tranh cãi. Một số nhà khoa học, đặc biệt là Matthew Wilkinson, cho rằng pteroid chĩa về phía trước, mở rộng màng. Quan điểm này mâu thuẫn với nghiên cứu năm 2007 của Chris Bennett, cho rằng pteroid không khớp như suy nghĩ trước đây và phải hướng vào trong cơ thể. Peters (2009) đề xuất rằng pteroid kết nối với phần 'yên' của động mạch quay và cả pteroid cùng khối xương cổ tay đã di chuyển về phía tâm. Quan điểm này được hỗ trợ bởi các mẫu vật của hai loài Changchengopterus pani và Darwinopterus linglongtaensis, cả hai đều cho thấy pteroid trong khớp nối với khối xương cổ tay gần.
Cổ tay thằn lằn bay gồm hai xương gần (gần cơ thể) và bốn xương xa (xa cơ thể), không tính xương pteroid, có thể nó là một xương cổ tay xa tiến hóa. Các xương cổ tay gần hợp thành một 'syncarpal' trong mẫu vật trưởng thành, trong khi ba trong số xương cổ tay xa kết hợp tạo thành một syncarpal xa. Xương cổ tay xa còn lại, gọi là carpal medial ('xương cổ tay trung gian'), cũng được gọi là xa bên, hoặc xương trước trục, khớp trên một mặt lồi thẳng đứng trên bề mặt trước của syncarpal xa. Các xương cổ tay trung gian có một cửa sổ lõm sâu mở ra phía trước, phần bụng và hơi trung gian, nơi pteroid khớp nối. Trong vài loài pterodactyloid như pteranodontian và azhdarchoid, xương bàn tay I-III nhỏ và không kết nối với cổ tay, thay vào đó tiếp xúc với xương bàn tay thứ tư, ở nyctosaurid các ngón trừ ngón cánh đã bị thoái hóa hoàn toàn.
Giới cổ sinh vật học đã tranh luận liệu màng cánh chính (brachiopatagia) có gắn vào chân sau không và nếu có thì ở đâu. Hóa thạch của các loài rhamphorhynchoid như Sordes, anurognathid Jeholopterus, và một pterodactyloid từ thành hệ Santana cho thấy màng cánh đã gắn vào chi sau, ít nhất là ở một số loài. Tuy nhiên, các loài dơi hiện đại và sóc bay cho thấy sự đa dạng đáng kể trong phạm vi màng cánh và có thể, như các nhóm này, các loài thằn lằn bay khác nhau có thiết kế cánh khác nhau. Thật vậy, phân tích tỷ lệ chiều dài chi của thằn lằn bay cho thấy sự biến động đáng kể, có thể phản ánh nhiều hình thái cánh khác nhau.
Nhiều loài thằn lằn bay, nếu không phải tất cả, đều có chân màng.
Đầu, răng và mào

Phần lớn hộp sọ thằn lằn bay có hàm dài với răng hình kim. Một số mẫu hóa thạch còn giữ được mô mỏ, dù vẫn có răng, nhưng mỏ nhỏ và chỉ ở đầu hàm, không liên quan đến răng. Một số loài mỏ cao cấp không có răng, như pteranodont và azhdarchid, có mỏ lớn và rộng hơn, giống như chim.
Khác với hầu hết các loài thằn lằn khác, các loài thằn lằn bay pterodactyloid có cửa sổ mũi và trước hố mắt gộp thành một lỗ duy nhất, gọi là cửa sổ mũi trước hố mắt. Đặc điểm này giúp làm nhẹ hộp sọ, phù hợp với hoạt động bay.
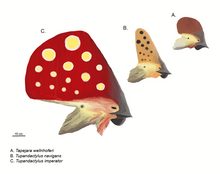
Một số loài thằn lằn bay có mào rất nổi bật. Điển hình và nổi tiếng nhất là mào chĩa sau của loài Pteranodon, nhưng một số loài khác như tapejarid và Nyctosaurus có mào cực lớn, thường kết hợp với mô sừng hoặc mô mềm mọc từ xương mỏ.
Từ những năm 1990, nhiều phát hiện và nghiên cứu kỹ lưỡng hơn đã cho thấy mào xuất hiện rộng rãi hơn ở nhiều loài so với dự đoán. Do mào thường kéo dài hoặc tạo từ keratin, một chất ít tạo hóa thạch hơn xương. Trường hợp của Pterorhynchus và Pterodactylus, chiều rộng thực sự của mào chỉ được phát hiện qua nhiếp ảnh tia cực tím. Phát hiện ra Pterorynchus và Austriadactylus, hai loài rhamphorhynchoid, cho thấy ngay cả thằn lằn bay nguyên thủy cũng có mào, trái với suy nghĩ trước đây rằng mào chỉ xuất hiện ở pterodactyloid tiến hóa.
Lông tơ (pycnofiber) của thằn lằn bay
Một số loài thằn lằn bay có lông giống tóc gọi là pycnofiber trên đầu và cơ thể, tương tự nhưng không cùng nguồn gốc với lông của động vật có vú. Năm 1831, Goldfuss lần đầu báo cáo một mảng thịt mờ từ mẫu Scaphognathus, và các khám phá cùng công nghệ mới đã chứng minh rằng thằn lằn bay có lớp lông pycnofiber. Dù không giống hệt lông của động vật có vú, pycnofiber là một cấu trúc độc đáo. Một số hóa thạch như Sordes pilosus và Jeholopterus ninchengensis có dấu ấn rõ ràng của pycnofiber trên đầu và cơ thể, tương tự quá trình tiến hóa hội tụ. Lớp sợi không bao phủ hàm lớn của thằn lằn bay trong hầu hết các mẫu vật được khám phá.
Một số nhà khoa học (Czerkas và Ji, 2002) cho rằng pycnofiber là tiền thân của lông vũ, nhưng dấu vết hóa thạch không giống như 'ống lông vũ' trên các loài khủng long giống chim. Pycnofiber có cấu trúc tương tự lông của khủng long chân thú, rất linh hoạt với sợi ngắn từ 5-7mm và đơn giản với một kênh trung tâm. 'Lông' thằn lằn bay bảo quản trong các thảm tập trung dày đặc, tương tự thảm quanh động vật có vú hóa thạch, cho thấy lớp lông dày như nhiều động vật có vú thời Đại Trung Sinh, ít nhất là trên các bộ phận phủ pycnofiber. Độ dày và diện tích bề mặt phủ pycnofiber chắc chắn đa dạng trong các loài thằn lằn bay.
Sự hiện diện của pycnofiber và nhu cầu bay lượn cho thấy thằn lằn bay có máu nóng. Việc thiếu pycnofiber trên cánh cho thấy lớp sợi không có chức năng khí động học, hỗ trợ giả thuyết rằng pycnofiber giúp điều hòa thân nhiệt, cần thiết để bảo tồn nhiệt từ hệ tiêu hóa của động vật máu nóng.
'Tóc' thằn lằn bay khác biệt rõ rệt với lông động vật có vú và sợi tơ của các loài khác, cần một tên gọi riêng. Thuật ngữ 'pycnofiber', nghĩa là 'sợi tơ dày đặc', được đề xuất trong một bài báo về dấu vết mô mềm của loài Jeholopterus bởi nhà cổ sinh vật học Alexander W.A. Kellner và đồng nghiệp năm 2009. Nghiên cứu mã di truyền của phôi cá sấu Mỹ cho thấy pycnofiber, vảy cá sấu và lông vũ tiến hóa tương đồng dựa trên cấu tạo keratin beta.
Lịch sử khai quật hóa thạch
Cổ sinh vật học
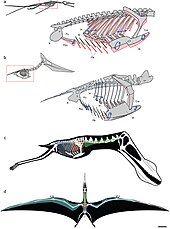

Khả năng bay
Phương thức bay lượn của các loài thằn lằn có cánh vẫn còn là điều bí ẩn và chưa thể mô phỏng chính xác.
Katsufumi Sato, một nhà khoa học Nhật Bản, đã sử dụng các loài chim hiện đại để tính toán và kết luận rằng một con dực long không thể bay. Trong cuốn sách Posture, Locomotion, and Paleoecology of Pterosaurs, ông giả thuyết rằng chúng bay được là do bầu không khí dày đặc và giàu oxy của cuối kỷ Phấn trắng. Tuy nhiên, cả Sato và các tác giả của cuốn sách này đều dựa trên những lý thuyết lỗi thời coi thằn lằn bay như chim biển, và kích thước không áp dụng cho các loài thằn lằn bay trên cạn như azhdarchid và tapejarid. Hơn nữa, Darren Naish kết luận rằng sự khác biệt về khí quyển giữa hiện tại và đại Trung Sinh không ảnh hưởng đến kích thước lớn của loài thằn lằn bay.
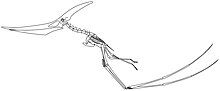
Một vấn đề khác là cách chúng cất cánh. Những giả thuyết trước đây cho rằng thằn lằn bay là động vật bay lượn máu lạnh, lấy nhiệt từ môi trường như các loài thằn lằn hiện đại, hơn là đốt cháy calo. Giả thuyết này bất hợp lý vì không rõ làm thế nào những con thằn lằn lớn như vậy có thể bay với sự trao đổi chất máu lạnh không hiệu quả, sử dụng cách cất cánh như chim chỉ bằng chân sau. Nghiên cứu sau này cho thấy chúng là loài máu nóng, có cơ bắp bay mạnh mẽ và sử dụng các chi trước để đi bộ. Mark Witton và Mike Habib cho rằng thằn lằn bay sử dụng cơ chế nhảy vọt để cất cánh. Sức mạnh của chi trước giúp chúng bay lên dễ dàng. Một khi đạt độ cao, chúng có thể bay với tốc độ lên đến 120 km/h và di chuyển hàng ngàn cây số.
Kích thước
Thằn lằn bay có nhiều kích cỡ khác nhau, với sải cánh từ 250 mm ở loài nhỏ nhất đến 10–11 m ở loài lớn nhất.
Túi khí và hệ hô hấp
Nghiên cứu năm 2009 cho thấy thằn lằn bay có hệ thống túi khí phổi và ống thở xương được điều khiển chính xác, hỗ trợ mô hình thông gió phổi chảy như ở chim. Hệ thống túi khí dưới da ở một số loài pterodactyloid giúp giảm khối lượng, làm chúng nhẹ hơn. Giống như cá sấu hiện đại, thằn lằn bay có túi bơm gan vì vành ngực vai của chúng không linh hoạt để di chuyển xương ức như chim và chúng có xương bụng khỏe. Do đó, hệ hô hấp của chúng có điểm tương đồng với cả hai nhánh thằn lằn chúa hiện đại.
Hệ thần kinh

Một nghiên cứu X-quang bề lõm hộp sọ đã chỉ ra rằng các loài Rhamphorhynchus muensteri và Anhanguera santanae có nhung não (flocculi) rất lớn. Thùy nhung tiểu não, nơi tích hợp các tín hiệu từ khớp, cơ bắp, da và các cơ quan thăng bằng, chiếm 7,5% tổng khối lượng não ở thằn lằn bay, vượt xa bất kỳ động vật có xương sống nào khác. Trong khi đó, chim có nhung não lớn hơn so với nhiều động vật khác, nhưng chỉ chiếm từ 1-2% tổng khối lượng não.
Nhung não truyền tín hiệu thần kinh nhỏ, tạo ra các cử động vô điều kiện ở cơ mắt, giúp ổn định hình ảnh trên võng mạc. Do kích thước sải cánh khổng lồ, thằn lằn bay cần xử lý nhiều thông tin cảm quan hơn, dẫn đến sự phát triển nhung não lớn. Sự khác biệt về khối lượng nhung não giữa thằn lằn bay và chim có thể do chim có bộ não lớn hơn nhiều; mặc dù điều này thường được coi là dấu hiệu cho thấy thằn lằn bay sống trong môi trường đơn giản hơn, nghiên cứu gần đây về cá sấu và bò sát khác chỉ ra rằng thằn lằn bay có mức độ thông minh cao với bộ não nhỏ. Nghiên cứu về Allkaruen cho thấy sự phát triển bộ não trong pterodactyloid là một quá trình mô-đun.
Di chuyển trên mặt đất
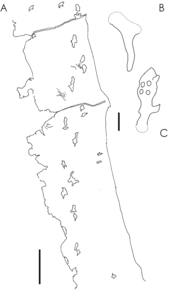
Lỗ ở hông thằn lằn bay hướng lên trên một chút và đầu xương đùi chỉ hướng vào trong một cách vừa phải, cho thấy thằn lằn bay có dáng đứng thẳng. Điều này cho phép chúng nâng đùi ngang trong khi bay, tương tự như hành vi của thằn lằn lượn.
Cuộc tranh luận diễn ra về việc thằn lằn bay di chuyển bằng bốn chân hay hai chân trên mặt đất. Vào những năm 1980, nhà cổ sinh vật học Kevin Padian đề xuất rằng các loài thằn lằn bay nhỏ hơn với chân dài, như chi Dimorphodon, có thể đi hoặc thậm chí chạy bằng hai chân, bên cạnh việc bay, giống như chi chẹo đất. Tuy nhiên, nhiều vết chân sau đó được phát hiện có dấu chân bốn ngón và dấu chân trước ba ngón độc đáo; đây là bằng chứng rõ ràng cho thấy thằn lằn bay đi bằng tất cả bốn chân.
Dấu chân hóa thạch cho thấy thằn lằn bay đặt toàn bộ gan bàn chân tiếp xúc với mặt đất, giống như nhiều động vật có vú như con người và gấu. Dấu chân từ azhdarchid và một số loài chưa xác định chỉ ra rằng thằn lằn bay khi di chuyển có bốn chân duỗi thẳng dưới cơ thể, một cách di chuyển hiệu quả năng lượng và phổ biến ở nhiều loài chim và động vật có vú hiện đại. Thực tế, bốn chi duỗi thẳng có thể đã xuất hiện ở tất cả các loài thằn lằn bay.

Mặc dù thằn lằn bay truyền thống được miêu tả là vụng về khi di chuyển trên mặt đất, nhưng giải phẫu của một số loài thằn lằn bay (đặc biệt là pterodactyloid) cho thấy chúng có khả năng đi bộ và chạy rất tốt. Các loài thằn lằn bay sớm từng được coi là cồng kềnh do các màng liên đùi lớn, nhưng giờ đây chúng được xem là có khả năng đi bộ hiệu quả trên mặt đất.
Xương chi trước của azhdarchid và ornithocheirid dài hơn bất thường so với các loài thằn lằn khác, và ở azhdarchid, xương cánh tay và bàn tay đặc biệt kéo dài. Tổng thể, chi trước của azhdarchid có cấu trúc tương tự như động vật có móng guốc chạy nhanh. Trong khi chi sau của chúng dài hơn hầu hết các loài thằn lằn, nó không được thiết kế cho tốc độ cao, nhưng cho phép một sải chân dài. Mặc dù azhdarchid có thể không chạy nhanh, chúng vẫn có khả năng di chuyển tương đối nhanh và tiết kiệm năng lượng.
Bơi
Dấu chân của các ctenochasmatoid cho thấy rằng thằn lằn bay này dùng chân sau để bơi. Nhìn chung, chúng có bàn chân sau lớn và cơ thể rộng, cho thấy khả năng bơi lội tốt hơn so với các loài thằn lằn bay khác. Ngược lại, các loài Pteranodontian có thể đã có những đặc điểm về xương cánh tay cho phép chúng di chuyển dưới nước bằng bốn chân, và một số như boreopterid có khả năng bắt mồi khi bơi, khác với khả năng săn mồi trên không như chim cốc biển. Các đặc điểm này cũng xuất hiện ở các loài thằn lằn bay sau này như azhdarchid, mà có thể dùng chi để thoát khỏi nước khi cần. Loài nyctosaurid Alcione có bằng chứng cho thấy khả năng lặn bằng cánh tương tự như ó biển hiện đại và chim nhiệt đới.
Thức ăn và hành vi ăn uống
Truyền thống cho rằng hầu hết các loài thằn lằn bay đều là những kẻ săn mồi cá trên mặt nước, quan điểm này vẫn phổ biến trong văn hóa đại chúng. Tuy nhiên, thực tế cho thấy nhiều loài thằn lằn bay thực sự là động vật ăn thịt trên cạn, ăn tạp hoặc ăn côn trùng.
Một trong những nhóm không bao giờ được cho là ăn cá là họ Anurognathidae; các loài này có thể sống về đêm, săn côn trùng trên không, quan điểm này vẫn được chấp nhận đến ngày nay. Với các khớp nối linh hoạt ở cánh tay, cánh tam giác rộng, mắt lớn và đuôi ngắn, những con thằn lằn này có đặc điểm tương tự như chim ăn thịt hoặc một số loài dơi ăn côn trùng hiện đại, cho phép chúng di chuyển nhanh nhẹn ở tốc độ thấp.
Dimorphodon từng được tưởng tượng là một loài tương tự như hải âu cổ rụt, nhưng với cấu trúc hàm và cách di chuyển đặc biệt, cùng khả năng bay không tốt, cho thấy nó có thể là một loài ăn thịt trên cạn hoặc sống nửa trên cây. Có vẻ như nó là một động vật ăn thịt nhỏ như động vật có vú, bò sát vảy, và côn trùng lớn.
Campylognathoides thường được coi là một loài ăn tạp hoặc động vật ăn thịt trên cạn của các động vật có xương sống nhỏ, nhờ vào bộ răng hàm chắc khỏe của nó. Xương cánh tay khổng lồ giống như của một con 'khỉ đột' và hình thái cánh cao như chim ưng cho thấy nó có thể đã phát triển khả năng bắt mồi bằng cánh.
Eudimorphodont có thể được phân loại thành hai nhóm chính: những loài có cánh dài và khỏe như Campylognathoides, và những loài có cánh dài mảnh mai hơn. Nhóm đầu tiên, bao gồm Carniadactylus và Eudimorphodon, là những động vật trên không nhanh nhẹn, có thể ăn côn trùng do kích thước nhỏ; Eudimorphodon đã được tìm thấy với cá trong dạ dày, nhưng răng của nó cho thấy chế độ ăn uống linh hoạt. Nhóm thứ hai, như Austriadactylus và Caviramus, có khả năng sống trên mặt đất/nửa trên cạn và có thể là những kẻ săn mồi cơ hội. Caviramus có lực cắn mạnh mẽ, thích nghi để nghiền nát thức ăn cứng. Tất cả các eudimorphodont đều có răng dạng hàm phát triển tốt, như răng của Caviramus và Eudimorphodon, cho phép nhai thức ăn, với Austriadactylus và Eudimorphodon sở hữu một cặp răng hàm phát triển thành răng nanh dài.
Rhamphorhynchid có thể được chia thành hai nhóm. Nhóm đầu tiên có cơ thể với mỏ dài, cánh dài và mảnh, răng giống như kim và hàm dài mỏng, đại diện bởi các loài như Rhamphorhynchus hoặc Dorygnathus, chuyên ăn cá. Nhóm còn lại có cấu trúc cơ thể 'khỏe mạnh', như Sericipterus, Scaphognathus và Harpactognathus, có hàm răng khỏe và cánh rộng nhưng ngắn hơn, là những kẻ săn mồi trên mặt đất/trên không, hoặc những kẻ ăn mồi cơ hội như loài quạ.
Wukongopterids như Darwinopterus trước đây được coi là kẻ săn mồi trên không; tuy nhiên, do thiếu cấu trúc hàm mạnh mẽ hoặc cơ bắp bay phát triển như của Campylognathoides hoặc rhamphorhynchids 'khỏe mạnh', chúng hiện tại được xem là động vật ăn côn trùng sống trên cây hoặc nửa dưới đất. Đặc biệt, Darwinopterus robustidens dường như chuyên ăn bọ cánh cứng.
Trong số các loài pterodactyloid, chế độ ăn uống rất đa dạng. Pteranodontia bao gồm nhiều nhóm ăn cá như ornithocheiran, boreopterid, pteranodontid và nyctosaurid. Có vẻ như có nhiều cách sống khác nhau: ornithocheiran và nyctosaurid có thể là những kẻ săn mồi trên không giống như các cốc biển hiện đại (trừ phương pháp lặn thả của Alcione), trong khi boreopterid có khả năng lặn trong môi trường nước ngọt như cá heo sông Hằng và sông Ấn, và pteranodont có khả năng lặn thả giống như chim điên và ó biển. Riêng istiodactylid dường như là loài ăn xác thối. Có bằng chứng cho thấy một số ornithocheirid săn mồi trên mặt đất.
Archaeopterodactyloidea bao gồm nhiều dực long kiếm ăn ở môi trường ven biển hoặc nước ngọt. Germanodactylus và Pterodactylus có thể là các loài ăn cá, trong khi Ctenochasmatidae lọc nước để ăn, với nhiều răng nhỏ giúp lọc các sinh vật nhỏ. Đặc biệt, Pterodaustro đã được ghi nhận là có cách ăn lọc giống như hồng hạc. Chế độ ăn của Cycnorhamphus vẫn là một bí ẩn; hàm cong lạ lùng của nó không rõ thuộc về chế độ ăn nào. Có thể nó ăn động vật có vỏ và sứa.
Ngược lại, Azhdarchoidea chủ yếu là những loài dực long sống trên cạn. Tapejarids được xem là động vật ăn tạp sống trên cây, ăn nhiều loại thực vật nhưng cũng có thể ăn cả côn trùng và động vật có xương sống nhỏ. Dsungaripterid được cho là ăn nhuyễn thể, sử dụng hàm mạnh mẽ để nghiền nát vỏ nhuyễn thể và động vật giáp xác. Dù quan điểm hiện tại là dsungaripterid là loài ăn tạp, chúng được cho là thích nghi với việc di chuyển trên mặt đất hơn là lội nước và được tìm thấy trong các khu vực lắng đọng đất liền. Thalassodromid có thể là động vật ăn thịt trên cạn. Thalassodromeus được đặt tên theo kiểu săn mồi gọi là 'ăn lướt' (khi một con chim bay sà trên mặt nước và dùng mỏ lướt trên bề mặt để bắt các động vật phù du hoặc cá), tuy nhiên nó (và các loài thằn lằn bay khác) không có khả năng này và thay vào đó là động vật ăn thịt săn mồi lớn, tương tự như phorusrhacidae. Đặc điểm này thể hiện qua các khớp hàm được gia cố và lực cắn mạnh.
Các loài azhdarchid nổi tiếng được biết đến như là những kẻ săn mồi trên mặt đất, tương tự như chim mỏ sừng hoặc một số loài cò, ăn bất kỳ con mồi nào mà chúng có thể nuốt. Hai ngoại lệ nổi bật là Hatzegopteryx, một loài săn mồi mạnh mẽ chuyên săn các loài động vật lớn, bao gồm cả khủng long cỡ trung bình; và Alanqa, có thể là loài ăn sinh vật nhuyễn thể.
Các lonchodectid có tỷ lệ cơ thể tương tự như azhdarchoid và có thể có tập tính tương tự. Tuy nhiên, chúng nổi bật với các răng khá đặc biệt, và loài Prejanopterus có phần hàm trên cong như cái móc câu, điều này có thể chỉ ra một số lối sống đặc biệt.
Loài săn mồi tự nhiên
Dực long được cho là đã trở thành con mồi của các loài khủng long chân thú. Trong số báo ngày 1 tháng 7 năm 2004 của tạp chí Nature, nhà cổ sinh vật học Eric Buffetaut mô tả một hóa thạch từ thời Phần Trắng sớm, bao gồm ba đốt sống cổ của một con thằn lằn bay và một số răng bị gãy của một con khủng long xương gai, có thể là Irritator, cắm vào con thằn lằn bay. Những đốt sống này không có dấu hiệu bị tiêu hóa, vì các khớp nối vẫn còn nguyên.
Sinh sản và tăng trưởng

Thông tin về sự sinh sản của dực long còn rất hạn chế, và trứng thằn lằn bay rất hiếm. Trứng dực long đầu tiên được phát hiện trong các mỏ đá ở Liêu Ninh, nơi cũng khai quật được những con khủng long lông vũ. Trứng bị nén nhưng không bị nứt, cho thấy chúng có vỏ da giống như trứng của các loài thằn lằn hiện đại. Một quả trứng thuộc chi Darwinopterus, mô tả vào năm 2011, cũng có vỏ da và kích thước nhỏ hơn so với kích thước của mẹ, tương tự như ở bò sát hiện đại nhưng khác với chim. Vào năm 2014, năm quả trứng không bị nén của Hamipterus tianshanensis được tìm thấy ở Tây Bắc Trung Quốc. Kiểm tra bằng kính hiển vi điện tử cho thấy lớp vỏ chứa calci mỏng và màng bên dưới. Một nghiên cứu năm 2007 cho thấy có khả năng dực long cũng chôn trứng như cá sấu và rùa hiện đại. Việc chôn trứng có thể giúp phát triển dực long bằng cách giảm trọng lượng, nhưng cũng đặt ra thách thức trong việc sống và sinh sản trong các môi trường cạnh tranh của loài chim.
Một mẫu của Darwinopterus cho thấy một số loài thằn lằn bay có tới hai buồng trứng chức năng, khác với chim chỉ có một buồng trứng chức năng. Điều này cho phép chúng duy trì khả năng bay bằng năng lượng hiệu quả hơn.
Màng cánh của phôi thằn lằn bay được bảo tồn tốt, cho thấy rằng chúng có khả năng cất cánh ngay sau khi sinh. Các hóa thạch của những con thằn lằn bay mới nở từ vài ngày đến một tuần tuổi (gọi là 'flaplings') đã được phát hiện, đại diện cho nhiều họ thằn lằn bay, bao gồm pterodactylids, rhamphorhinchids, ctenochasmatids và azhdarchids. Các xương của chúng cho thấy độ cứng cáp cao và tỷ lệ cánh tương tự như người trưởng thành. Trên thực tế, nhiều cá thể non từng được coi là loài riêng biệt trong quá khứ. Con non thường được tìm thấy cùng với cá thể trưởng thành và vị thành niên của cùng một loài, chẳng hạn như flaplings của Pterodactylus và Rhamphorhynchus trong tầng đá vôi Solnhofen của Đức, và flaplings của pterodaustro từ Brazil. Tất cả được phát hiện trong môi trường nước sâu xa bờ.
Mặc dù chưa rõ liệu các loài thằn lằn bay có chăm sóc con non hay không, nhưng khả năng bay ngay sau khi chào đời và sự xuất hiện của nhiều cá thể non ở xa tổ và bên cạnh những con trưởng thành đã khiến nhiều chuyên gia, như Christopher Bennett và David Unwin, kết luận rằng con non chỉ phụ thuộc vào bố mẹ trong một khoảng thời gian ngắn để phát triển cánh đủ lớn để bay, và sau đó tự rời tổ chỉ trong vài ngày sau khi nở. Chúng có thể đã dựa vào chất dinh dưỡng từ lòng đỏ trứng để nuôi cơ thể trong vài ngày đầu đời, giống như các loài bò sát hiện đại, thay vì phụ thuộc vào cha mẹ để tìm kiếm thức ăn.
Tốc độ phát triển của dực long sau khi nở có sự khác biệt rõ rệt giữa các nhóm. Các loài thằn lằn bay sơ khai với đuôi dài hơn, như Rhamphorhynchus, có tốc độ tăng trưởng trung bình từ 130% đến 173% trong những năm đầu đời, nhanh hơn một chút so với cá sấu. Sự phát triển của chúng chậm lại sau khi đạt đến trưởng thành tình dục, và Rhamphorhynchus cần hơn ba năm để đạt kích thước tối đa. Ngược lại, các loài thằn lằn bay tiên tiến thuộc họ pterodactyloid lớn hơn, như Pteranodon, hoàn thành việc phát triển kích thước hoàn chỉnh trong năm đầu đời. Những pterodactyloids này có sự tăng trưởng xác định, nghĩa là chúng đạt kích thước lớn nhất và ngừng phát triển từ đó.
Tập tính hằng ngày
So sánh giữa các vòng xơ cứng của mắt ở thằn lằn bay và chim cùng các loài bò sát hiện đại đã giúp tái dựng lại thói quen sinh hoạt của thằn lằn bay. Các loài dực long như Pterodactylus, Scaphognathus, và Tupuxuara được cho là hoạt động vào ban ngày, trong khi Ctenochasma, Pterodaustro, và Rhamphorhynchus có thể là loài hoạt động về đêm, còn Tapejara có thói quen hoạt động ngắn hạn trong suốt cả ngày. Do đó, Ctenochasma và Rhamphorhynchus có thể ăn cá tương tự như các loài chim biển hiện đại, và Pterodaustro có thể lọc thức ăn giống như các ngỗng hiện đại hoạt động về đêm. Sự khác biệt trong thói quen hoạt động của các loài tại vùng Solnhofen như Ctenochasma, Rhamphorhynchus, Scaphognathus, và Pterodactylus có thể chỉ ra phân vùng sinh thái đặc trưng giữa các chi này.
Phân loại
Tổ tiên

Do dực long đã tiến hóa mạnh mẽ để bay lượn và các hóa thạch chuyển tiếp chưa được mô tả, tổ tiên của chúng vẫn chưa rõ ràng. Các dực long cổ nhất đã hoàn toàn thích ứng với cuộc sống trên không. Từ thời Seeley, người ta cho rằng dực long có thể xuất phát từ 'các thằn lằn chúa' hay archosauromorpha. Vào những năm 1980, phân tích cho thấy chúng thuộc nhánh avemetatarsalia (các thằn lằn chúa gần với khủng long hơn cá sấu). Kevin Padian coi khám phá này làm vững chắc giả thuyết của ông rằng dực long là động vật máu nóng đi bằng hai chân. Tuy nhiên, các phân tích thời kỳ đầu dựa trên số lượng phân loại và các đặc điểm hạn chế nên kết luận không chắc chắn. Một số nhà nghiên cứu quan trọng đã bác bỏ kết luận của Padian và đưa ra các giả thuyết khác. David Unwin đề xuất tổ tiên chung thuộc nhóm archosauromorpha cơ bản (các loài cổ hơn), như chi Tanystropheus. Một tổ tiên thuộc nhóm archosauriform cơ bản, như chi Euparkeria, cũng được đề xuất. Một số archosauromorph cơ bản, như chi Sharovipteryx, với màng da ở chi sau giúp lướt trên không, được xem là ứng cử viên sáng giá. Năm 1999, Michael Benton phát hiện dực long thuộc avemetatarsalia có quan hệ gần với chi Scleromochlus, và đặt tên cho nhóm này là Ornithodira để bao gồm dực long và khủng long.

Chris Bennett (1996) và David Peters (2000) đã chỉ ra rằng dực long có thể là protosauria hoặc có mối quan hệ gần gũi với nhóm này. Peters đã áp dụng kỹ thuật DGS, sử dụng phần mềm chỉnh sửa ảnh để phân tích hóa thạch dực long. Nếu Peters loại bỏ các đặc điểm phần chi sau khỏi phân tích, dực long có thể được coi là họ hàng gần của protosauria, nhằm kiểm tra giả thuyết rằng các đặc điểm này là kết quả của sự tiến hóa hội tụ với khủng long. Tuy nhiên, phân tích sau đó của Dave Hone và Michael Benton (2007) không thể tái tạo kết quả của Peters. Hone và Benton phát hiện dực long gần gũi với khủng long, ngay cả khi không xét đến phần chi sau, và chỉ trích các nghiên cứu của Peters vì thiếu bằng chứng hóa thạch trực tiếp. Họ kết luận rằng mặc dù cần nghiên cứu thêm về các pterosauromorph cơ bản, dực long hiện được coi là một nhánh của avemetatarsalia. Trong phân tích của họ, dực long là nhóm chị em của Scleromochlus hoặc nằm giữa nó và Lagosuchus trên cây tiến hóa nhánh ornithodira. Một phân tích năm 2011 của Sterling Nesbitt với dữ liệu phong phú hơn hỗ trợ mạnh mẽ cho giả thuyết dực long là avemetatarsalia, mặc dù chi Scleromochlus không được đưa vào nghiên cứu do tình trạng bảo quản kém. Một nghiên cứu năm 2016 của Martin Ezcurra về archosauromorph, bao gồm các họ hàng dực long khác, cũng kết luận rằng dực long gần khủng long hơn và không liên quan đến các đơn vị phân loại cơ bản hơn. Bennett, trong nghiên cứu năm 2020 của mình, lập luận rằng cả Scleromochlus và dực long đều là archosauromorph không phải thằn lằn chúa, nhưng không có mối quan hệ gần gũi. Một nghiên cứu năm 2020 sau đó đề xuất rằng nhánh lagerpetidae của thằn lằn chúa là nhánh chị em của pterosauria, dựa trên các hóa thạch hộp sọ và chi trước mới mô tả cho thấy nhiều điểm tương đồng với dực long, cùng với bản phục dựng não lagerpetidae cho thấy điểm tương đồng về thần kinh.
Một vấn đề quan trọng là nguồn gốc của khả năng bay lượn. Giống như chim, có hai giả thuyết chính: 'từ đất lên' hoặc 'từ cây xuống'. Giả thuyết leo lên cây cho rằng việc leo lên cao và trọng lực đã cung cấp năng lượng và áp lực chọn lọc cho khả năng bay. Rupert Wild (1983) đã đề xuất một loài giả thuyết 'propterosaurus', một thằn lằn sống trên cây với màng giữa các chi giúp sà xuống an toàn và dần dần phát triển ngón tay thứ tư dài ra để lướt. Tuy nhiên, các nghiên cứu sau đó không phù hợp với mô hình này. Cả protorosaur và ornithodiran không tương đương với thằn lằn về mặt sinh học. Hơn nữa, sự chuyển tiếp từ bay lướt sang bay vỗ vẫn chưa rõ. Nghiên cứu gần đây về chi sau của dực long cho thấy mối liên hệ với Scleromochlus. Giống như chi thằn lằn chúa này, các dực long cơ bản có chi sau đi bằng cả gan bàn chân (plantigrade), cho thấy thích ứng để nhảy vọt.
Tuyệt chủng

Trước đây, người ta cho rằng sự cạnh tranh với các loài chim thời kỳ đầu có thể đã dẫn đến sự tuyệt chủng của nhiều loài dực long. Một phần lý do là do các nhà khoa học tin rằng vào cuối kỷ Phấn trắng, chỉ còn lại các loài dực long lớn, điều này đã được chứng minh là không chính xác. Các loài dực long nhỏ hơn được cho là đã tuyệt chủng và bị thay thế bởi các loài chim. Tuy nhiên, sự suy giảm của dực long (nếu có) không có liên quan rõ ràng đến sự phát triển của các loài chim, vì sự chồng chéo sinh thái giữa hai nhóm này không đáng kể. Thực tế, một số môi trường sinh thái của chim đã được dực long chiếm lĩnh trước sự kiện tuyệt chủng Phấn Trắng-Cổ Cận. Cuối kỷ Phấn trắng, sự kiện KT đã xóa sổ tất cả các loài khủng long không phải là chim cùng với nhiều loài động vật khác, bao gồm cả dực long.
Vào đầu những năm 2010, một số đơn vị phân loại dực long mới được phát hiện thuộc tầng Campanian/Maastrichtian, như chi ornithocheirid Piksi và 'Ornithocheirus', pteranodontid và nyctosaurid, nhiều chi tapejarid và một chi không phải azhdarchid chưa xác định là Navajodactylus. Các loài dực long azhdarchoid nhỏ cũng xuất hiện ở tầng Campanian. Điều này cho thấy rằng quần xã dực long vào cuối kỷ Phấn trắng đa dạng hơn nhiều so với những gì đã biết trước đây, thậm chí có thể không suy giảm đáng kể so với kỷ Phấn trắng sớm.
Các loài dực long nhỏ kích thước đã được tìm thấy trong thành hệ Csehbánya. Những phát hiện gần đây về một con azhdarchid trưởng thành có kích thước bằng con mèo cho thấy rằng dực long nhỏ từ kỷ Phấn trắng muộn hiếm khi được bảo tồn trong các lớp đá.
Phát sinh chủng loại học
Trong phân loại phát sinh loài, nhánh pterosauria thường được xác định dựa trên các điểm nút và liên kết với những đơn vị phân loại đã được nghiên cứu kỹ lưỡng cùng các loài được coi là nguyên thủy. Một nghiên cứu năm 2003 đã định nghĩa Pterosauria là 'Tổ tiên chung gần đây nhất của Anurognathidae, Preondactylus và Quetzalcoatlus cùng tất cả các hậu duệ của chúng.' Tuy nhiên, định nghĩa này có thể bỏ sót các loài nguyên thủy hơn ngoài Pterosauria. Để khắc phục điều này, một định nghĩa mới đã được đưa ra, tập trung vào đặc điểm giải phẫu thay vì các loài cụ thể, đó là ngón tay thứ tư dài bất thường hỗ trợ màng cánh. Định nghĩa 'dựa trên đặc điểm apomorphy' này đã được PhyloCode thông qua vào năm 2020, 'Ngành đặc trưng bởi apomorphy ngón tay thứ tư phì đại để hỗ trợ màng cánh, kế thừa bởi loài Pterodactylus (trước đây là Ornithocephalus) antiquus (Sömmerring 1812).' Một nhánh rộng hơn, tên là Pterosauromorpha, được định nghĩa là tất cả các ornithodiran gần gũi hơn với dực long so với khủng long.
Việc phân loại các loài dực long đã gặp khó khăn trong lịch sử do sự thiếu hụt thông tin từ các hóa thạch. Bước sang thế kỷ 21, những khám phá mới đang dần lấp đầy những khoảng trống này và tạo ra cái nhìn toàn diện hơn về sự tiến hóa của dực long. Truyền thống chia dực long thành hai cận bộ: Rhamphorhynchoidea, dực long 'nguyên thủy' với đuôi dài, và Pterodactyloidea, dực long 'tiến hóa' với đuôi ngắn. Tuy nhiên, phân loại này đã trở nên lạc hậu. Rhamphorhynchoidea là một nhóm cận ngành vì các pterodactyloid tiến hóa trực tiếp từ chúng, không phải từ cùng tổ tiên chung. Do đó, với sự áp dụng ngày càng rộng rãi của mô tả theo nhánh học, phương pháp truyền thống đã không còn được ưu chuộng bởi các nhà khoa học.
Mối quan hệ chính xác giữa các loài dực long vẫn chưa được xác định rõ ràng. Nhiều nghiên cứu trước đây về mối quan hệ của dực long có dữ liệu hạn chế và không nhất quán. Tuy nhiên, các nghiên cứu mới với cơ sở dữ liệu lớn hơn đang bắt đầu làm sáng tỏ vấn đề. Biểu đồ nhánh (cây phát sinh chủng loại) dưới đây dựa trên phân tích của Longrich, Martill và Andres năm 2018, với tên nhánh được đặt bởi Andres et al (2014).










| Pterosauria |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||