
| Đây là một trong loạt bài về |
| Hóa sinh |
|---|
| Các nét quan trọng |
|
|
| Lịch sử và chủ đề |
|
|
|
|
| Thuật ngữ |
|
| Lĩnh vực: Sinh học, Sinh học phân tử, tế bào |
Sinh học phân tử là một phân ngành sinh học nhằm nghiên cứu cơ sở phân tử của các hoạt động sinh học bên trong và giữa các tế bào, bao gồm quá trình tổng hợp, biến đổi, cơ chế, và tương tác phân tử. Nghiên cứu cấu trúc hóa học và vật lý của các đại phân tử được gọi là sinh học phân tử.
Sinh học phân tử lần đầu tiên được mô tả là một phương pháp tiếp cận tập trung vào nền tảng của các hiện tượng sinh học, nhằm khám phá cấu trúc và các tương tác của những phân tử sinh học, và cách những tương tác này giải thích các hiện tượng của sinh học.

Năm 1945, thuật ngữ 'sinh học phân tử' được sử dụng bởi nhà vật lý William Astbury. Sự phát triển trong lĩnh vực sinh học phân tử xảy ra khá muộn do cơ thể sinh vật là một hệ thống phức tạp, và cách tiếp cận đơn giản và thuận tiện nhất là sử dụng các vi khuẩn và thể thực khuẩn chỉ có thể đưa ra thông tin về quá trình sinh học cơ bản. Vào năm 1953, hai nhà khoa học trẻ lúc bấy giờ tên Francis Crick và James Watson làm việc tại đơn vị Hội đồng Nghiên cứu Y khoa, phòng thí nghiệm Cavendish, Cambridge (nay là Phòng thí nghiệm Sinh học Phân tử MRC), đã tạo ra một mô hình mạch xoắn kép của ADN. Mô hình mới này đã thay đổi toàn bộ cấu trúc ADN mà họ đã đề xuất trước đó dựa trên các nghiên cứu được thực hiện bởi Rosalind Franklin và Maurice Wilkins. Sau mô hình cấu trúc DNA, các nghiên cứu của hai nhà khoa học tập trung chủ yếu vào mục tiêu tìm ra DNA bên trong các vi sinh vật, thực vật, và động vật khác.
Sinh học phân tử không chỉ đơn giản là nghiên cứu các phân tử và sự tương tác của chúng. Nói chính xác hơn, sinh học phân tử là tập hợp các kỹ thuật được phát triển kể từ khi lĩnh vực này mới bắt đầu, cho phép các nhà khoa học tìm hiểu về các quá trình phân tử. Một kỹ thuật đáng chú ý đã cách mạng hóa lĩnh vực này là phản ứng chuỗi polymerase (tiếng Anh: Polymerase Chain Reaction hay PCR), được phát triển vào năm 1983. PCR là một phản ứng khuếch đại một lượng nhỏ ADN, và được sử dụng trong nhiều trong các ngành khoa học.
Luận thuyết trung tâm của sinh học phân tử cho rằng dòng thông tin chạy theo một chiều từ DNA đến mRNA rồi đến protein. Quá trình phiên mã, dịch mã và sao chép DNA tuân theo quy luật này. Một ngoại lệ phát hiện thấy ở một số virus mang RNA, có quá trình gọi là phiên mã ngược, tạo ra một bản sao DNA từ RNA hệ gen của chúng.
Sinh học phân tử đóng một vai trò quan trọng trong sự hiểu biết về cấu trúc, chức năng, và quá trình kiểm soát nội môi bên trong các tế bào. Tất cả những tri thức trên đều có thể được sử dụng để phát triển các loại thuốc và phuơng pháp chẩn đoán bệnh mới, và hiểu rõ hơn về các quá trình sinh lý của tế bào. Một số nghiên cứu lâm sàng và liệu pháp y tế phát triển từ sinh học phân tử được sử dụng trong liệu pháp gen. Trong khi đó, việc sử dụng sinh học phân tử hoặc sinh học tế bào phân tử trong y học hiện nay được gọi là y học phân tử.
Lịch sử
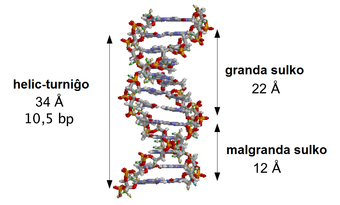
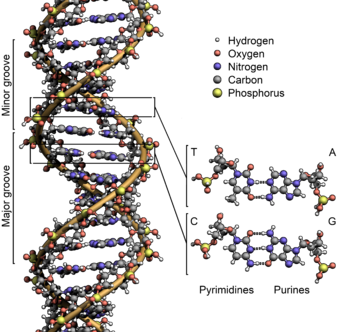
Sinh học phân tử là sự giao thoa giữa ngành di truyền học và hóa sinh. Ở thế kỷ 20 khi di truyền học và hóa sinh vừa hình thành và bắt đầu phát triển, cả hai lĩnh vực rõ ràng đều nỗ lực tìm cách xác định các cơ chế phân tử giúp tạo nên nền tảng cho các chức năng quan trọng của tế bào. Những tiến bộ trong sinh học phân tử có liên quan chặt chẽ đến sự phát triển của các công nghệ mới và sự tối ưu hóa trong cả hai lĩnh vực di truyền học và hóa sinh. Sinh học phân tử đã được làm sáng tỏ bởi công trình nghiên cứu của nhiều nhà di truyền học và hóa sinh, và do đó lịch sử của phân ngành này phụ thuộc vào sự hiểu biết của các nhà khoa học này và những thí nghiệm của họ.

Có thể nói, tất cả kiến thức hiện tại của sinh học phân tử đều bắt đầu từ hiện tượng biến nạp ở vi khuẩn. Năm 1928, Frederick Griffith, đã quan sát thấy hiện tượng biến đổi từ vi khuẩn này sang vi khuẩn khác mà ngày nay được gọi là biến nạp, và ở thời điểm đó, ông không thể tìm ra lý do để giải thích sự việc này. Do đó, người ta còn gọi ông là nhà vi sinh vật học. Mãi cho đến năm 1944, ba nhà khoa học Oswald Avery, Colin Macleod và Maclyn McCarty, đã chứng minh toàn bộ hiện tượng biến nạp ở vi khuẩn. Vào năm 1930, hai năm sau thí nghiệm của Griffith, sinh học phân tử được thành lập như một phân ngành khoa học chính thức. Tuy nhiên, thuật ngữ “sinh học phân tử” vẫn chưa tồn tại cho đến năm 1938 khi nhà khoa học Warren Weaver, người đang làm việc với tư cách là giám đốc lĩnh vực Khoa học Tự nhiên tại Quỹ Rockefeller, đặt tên phân ngành này là sinh học phân tử.
Sau một loạt các thí nghiệm được trải dài sang nhiều năm, những nhà nghiên cứu sinh học phân tử mới kết luận được rằng ADN là vật chất di truyền cơ bản gây ra các biến đổi di truyền. Thành phần cơ bản của ADN chứa bốn base là - Adenin (A), Guanin (G), Thymine (T), và Cytosine (C). Dựa trên thành phần hóa học và thông tin từ tinh thể học tia X được thực hiện bởi Maurice Wilkins và Rosalind Franklin, cấu trúc ADN đã được đề xuất bởi James Watson và Francis Crick. Tuy nhiên, trước khi Watson và Crick đề xuất cấu trúc ADN, vào năm 1950, nhà khoa học người Áo Erwin Chargaff, đã đề xuất một lý thuyết hay quy luật mà ngày nay được gọi là quy luật Chargaff. Quy luật nêu rằng tỷ lệ của Adenin với Thymine, và Guanin với Cytosine là bằng nhau
- Quy luật Chargaff:
Nguyên văn:
'Chargaff's rule stated that DNA from any species of any organism should have a 1:1 stoichiometric ratio of purine and pyrimidines (i.e., A+G=T+C) and, more specifically, that the amount of guanine should be equal to cytosine and the amount of adenine should be equal to thymine. This pattern is found in both strands of the DNA'.
Tạm dịch:
'Quy luật Chargaff nêu rằng ADN từ bất kỳ loài sinh vật nào đều phải có tỷ lệ phân cực là 1:1 giữa purin và pyrimidin (tức là A + G = T + C) và đặc biệt hơn, số lượng guanin phải bằng cytosine, và số lượng adenin phải bằng thymine. Mô típ này được tìm thấy ở cả hai mạch của ADN'.
Lĩnh vực di truyền học ra đời như một nỗ lực nhằm tìm hiểu các cơ chế phân tử của di truyền gen và cấu trúc của một gen. Gregor Mendel đã đi tiên phong trong công việc này vào năm 1866 khi ông lần đầu tiên viết các quy luật di truyền dựa trên các nghiên cứu của mình về phép lai giao phối ở cây đậu. Một trong những quy luật di truyền là quy luật phân li. Quy luật này phát biểu rằng các cá thể lưỡng bội có hai alen cho một gen cụ thể sẽ truyền một trong các alen này cho con cái của chúng. Do có vai trò quan trọng trong lĩnh vực di truyền học, các nghiên cứu về di truyền gen thường được gọi là di truyền Mendel.
Một cột mốc quan trọng trong phân ngành sinh học phân tử là việc tìm ra cấu trúc của ADN. Công việc này bắt đầu vào năm 1869 bởi Friedrich Miescher, một nhà hóa sinh người Thụy Sĩ, người đầu tiên đề xuất một cấu trúc gọi là nuclein, mà ngày nay chúng ta biết là axit deoxyribonucleic, hay DNA. Ông đã phát hiện ra chất hóa học này bằng cách nghiên cứu các thành phần có trong mủ dính trên băng dán cá nhân, và ghi nhận các đặc tính độc đáo của 'chất chứa phosphor'. Một người đóng góp quan trọng khác cho mô hình ADN là Phoebus Levene, người đã đề xuất 'mô hình polynucleotide' của ADN vào năm 1919 dựa trên kết quả của các thí nghiệm sinh hóa của ông trên nấm men. Vào năm 1950, Erwin Chargaff đã mở rộng nghiên cứu của Levene và làm sáng tỏ một số đặc tính quan trọng của axit nucleic. Thứ nhất, trình tự của axit nucleic trên các loài khác nhau. Thứ hai, tổng nồng độ của purin (A và G) luôn bằng tổng nồng độ của pyrimidin (C và T), và điều này bây giờ được gọi là quy luật Chargaff. Năm 1953, James Watson và Francis Crick công bố cấu trúc xoắn kép của ADN dựa trên công trình tinh thể học tia X do Rosalind Franklin và Maurice Wilkins thực hiện. Watson và Crick mô tả cấu trúc của ADN và phỏng đoán về ảnh hưởng của cấu trúc độc đáo này đối với cơ chế nhân bản ADN. Sau cùng, J. D. Watson và F. H. C. Crick đã được trao giải Nobel năm 1962, cùng với Maurice Wilkins, vì đã đề xuất một mô hình về cấu trúc của ADN.
Thời gian trôi qua, vào năm 1964, Kjeld Marker và Frederick Sanger đã phát hiện ra một ARN vận chuyển amioacyl đặc biệt ở E.coli, được gọi là ARN vận chuyển N-formyl-methionyl và giải thích rằng phân tử này đóng một vai trò trong cơ chế đặc biệt của quá trình kéo dài mạch khi tế bào dịch mã. Ông đã được trao giải Nobel thứ hai vì đã tìm ra trình tự hoàn chỉnh bao gồm 5.400 nucleotide trên mạch đơn ADN của thể thực khuẩn Phi X174.
Vào năm 1961, các nhà khoa học đã chứng minh được rằng khi một gen mã hóa thành protein, ba base liên tiếp của ADN của gen sẽ chỉ định từng axit amin kế tiếp của protein. Do đó, mã di truyền là mã bộ ba, trong đó mỗi bộ ba (được gọi là codon) chỉ định một axit amin cụ thể. Hơn nữa, các nghiên cứu chỉ ra rằng các codon không chồng chéo với nhau trong trình tự ADN và mỗi trình tự được đọc từ một điểm bắt đầu cố định.
Trong giai đoạn 1962–1964, thông qua việc sử dụng các thể thực khuẩn mang đột biến gây tự chết có điều kiện, các nhà sinh học phân tử đã đạt được những tiến bộ cơ bản trong việc hiểu biết về các chức năng và tương tác của các protein được sử dụng trong quá trình sao chép, sửa chữa, và tái tổ hợp ADN, và trong việc hình thành các cấu trúc phân tử.
Thí nghiệm Griffith
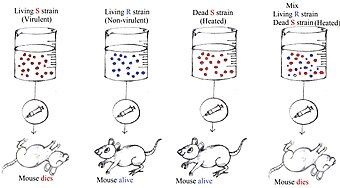
Năm 1928, Frederick Griffith, đã phát hiện độc tính của phế cầu khuẩn khi loại vi trùng này khiến chuột trong phòng thí nghiệm tử vong. Dựa trên di truyền Mendel, tử tưởng được dùng phổ biến vào lúc bấy giờ, việc chuyển gen chỉ có thể xảy ra từ tế bào mẹ sang tế bào con. Griffith đưa ra một lý thuyết khác với lý luận rằng sự chuyển gen cũng có thể xảy ra giữa các thành viên của cùng một thế hệ, và quá trình này được gọi là chuyển gen ngang (tiếng Anh: Horizontal Gene Transfer hay HGT). Ngày nay, hiện tượng này được gọi là biến nạp.
Trong thí nghiệm của mình, Griffith đã đề cập đến loài phế cầu khuẩn Streptococcus pneumoniae. Loài vi khuẩn này có hai chủng khác nhau: một loại có độc tính và bờ nhẵn smooth (S), và một loại trung tính không độc và nhám rough (R). Nhìn bằng mắt thường, dòng S có vẻ ngoài lấp lánh do sự hiện diện của một loại polysaccharide đặc trưng ở vỏ bọc tạo thành bởi polyme của axit glucuronic và glucose. Nhờ sự hiện diện của lớp polysaccharide, hệ thống miễn dịch của vật chủ không thể phát hiện vi khuẩn, và điều này giúp sinh vật này có thể phát độc và giết chết vật chủ. Trong khi đó, loài trung tính không độc R còn lại, do thiếu vỏ bọc chứa lớp polysaccharide, nên loài này có vẻ ngoài sần sùi và xỉn màu.
Sự hiện diện của vỏ bọc ở phế cầu khuẩn có thể được xác định thông qua di truyền học. Chủng S và R có thể tồn tại ở nhiều dạng khác nhau như S-I, S-II, S-III, v.v,. và tương ứng là R-I, R-II, R-III, v.v. Tất cả các phân nhóm của chủng S và R nêu trên được phân loại dựa vào loại kháng nguyên khác nhau mà chúng tạo ra.
Thí nghiệm Hershey-Chase
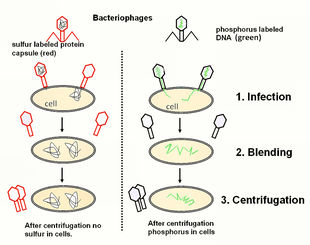
Thí nghiệm của Hershey và Chase đã xác nhận ADN là vật chất di truyền khiến chuột của trong thí nghiệm Griffith bị nhiễm độc của phế cầu khuẩn và tử vong. Thí nghiệm này còn được gọi là thí nghiệm máy xay sinh tố do hai nhà khoa học liên tục dùng loại xay sinh tố thường được sử dụng nhà bếp như một thiết bị chính. Trong thí nghiệm của mình, Hershey và Chase sử dụng E.coli và thể thực khuẩn, và kết quả đã chứng minh rằng ADN của thể thực khuẩn, sau khi lây nhiễm vào tế bào vi khuẩn, có chứa tất cả thông tin cần thiết để tổng hợp các thể thực khuẩn đời con cháu. Hai nhà khoa học đã đánh dấu lớp vỏ protein của thể thực khuẩn bằng đồng vị phóng xạ lưu huỳnh. Trong khi đó, ADN của thể thực khuẩn được đánh dẫu bằng đồng vị phóng xạ phốt pho. Sau đó, hai mẫu thể thực khuẩn tách biệt lần lượt được cho vào hai ống nghiệm khác nhau. Sau khi trộn thể thực khuẩn bị nhiễm phóng xạ và E.coli ở trong ống nghiệm, hai mẫu được ủ ở điều kiện thích hợp nhằm khiến cho thể thực khuẩn có thể chuyển vật chất di truyền (protein hoặc ADN) sang tế bào E.coli. Tiếp theo, hỗn hợp vi khuẩn và thể thực khuẩn được đưa vào máy xay sinh tố để trộn hoặc khuấy động nhằm tách thể thực khuẩn ra khỏi tế bào E.coli. Toàn bộ hỗn hợp được ly tâm. Phần cặn bên dưới đáy ống nghiệm có chứa tế bào E.coli được giữ lại và kiểm tra, còn phần loại chất lỏng được loại bỏ. Sau khi kiểm tra tế bào E.coli chỉ chứa đồng vị phóng xạ phốt pho, điều này cho thấy vật chất di truyền được biến nạp là ADN chứ không phải là protein.
ADN của thể thực khuẩn sau khi biến nạp sẽ được tích hợp vào ADN của E.coli. Do đó, phần ADN của E.coli bị nhiễm phóng xạ thực chất là của thể thực khuẩn. Khi vi khuẩn tiếp tục phát triển và sinh sản, đoạn ADN đột biến này có thể được truyền lại cho thế hệ tiếp theo, và từ đây, lý thuyết về tải nạp gen đã ra đời. Tải nạp là một quá trình trong đó ADN của vi khuẩn có chứa một đoạn ADN của thể thực khuẩn, và đoạn ADN này sẽ được truyền cho thế hệ vi khuẩn tiếp theo. Đồng thời, tải nạp có thể được xem là một ví dụ của hoạt động chuyển gen ngang.
Sinh học phân tử hiện đại
Khi xã hội loài người đang dần bước đến giữa những năm 20 của thế kỷ 21, sinh học phân tử cũng đang bước vào thời kỳ hoàng kim nhờ vào sự phát triển của khoa học kỹ thuật. Các công nghệ mới đang cho phép theo dõi các quá trình sinh học ở cấp độ nguyên tử ở thời gian thực. Ngày nay, các nhà sinh học phân tử có thể truy cập vào kho dữ liệu trình tự DNA của các loài đồ sộ với một mức chi phí hợp lý, và điều này đã tạo điều kiện phát triển các phương pháp chỉnh sửa gen mới ở các sinh vật khác bên cạnh sinh vật mô hình. Tương tự như vậy, các nhà sinh học phân tử tổng hợp sẽ thúc đẩy quá trình sản xuất công nghiệp các tiểu và đại phân tử thông qua việc phát triển các con đường trao đổi chất ngoại sinh trong các dòng tế bào nhân sơ và nhân thực khác nhau.
Dữ liệu trình tự sinh học ngày càng có giá cả phải chăng hơn khiến cho nó sử dụng trong nhiều lĩnh vực khoa học khác nhau. Điều này hứa hẹn sẽ thúc đẩy sự phát triển của các ngành công nghiệp ở các quốc gia đang phát triển, và giúp tăng khả năng các nhà nghiên cứu tư nhân có thể tiếp cận được thông tin cần thiết cho những thực nghiệm của học. Ngoài ra, các thí nghiệm chỉnh sửa gen dùng công nghệ CRISPR-Cas9 hiện có thể được thực hiện trên các sinh vật mới bởi các công ty tư nhân với giá dưới 10.000 đô la, và điều này sẽ giúp thúc đẩy cho sự phát triển của việc ứng dụng công nghệ này vào các lĩnh vực công nghiệp và y tế.
Quan hệ với các phân ngành sinh học khác
- Xét về định nghĩa:
- Sinh học phân tử là nghiên cứu về cơ sở phân tử của các hiện tượng sinh học, tập trung vào những quá trình tổng hợp, biến đổi, cơ chế, hay tương tác phân tử.
- Hóa sinh là nghiên cứu về các chất hóa học và các quá trình quan trọng xảy ra trong cơ thể sinh vật. Các nhà hóa sinh tập trung chủ yếu vào vai trò, chức năng, và cấu trúc của các đại phân tử như protein, lipid, cacbohydrat, và axit nucleic.
- Di truyền học là nghiên cứu về cách những khác biệt về mặt di truyền ảnh hưởng đến các sinh vật. Di truyền học cố gắng dự đoán cách các đột biến, các gen riêng lẻ, và các tương tác di truyền có thể ảnh hưởng đến biểu hiện gen và quá trình tạo nên một kiểu hình.
Khi các nhà khoa học thực hành những kỹ thuật cụ thể cho nghiên cứu sinh học phân tử, họ sẽ thường kết hợp các kỹ thuật này với nhiều phương pháp khác từ phân ngành di truyền học và hóa sinh. Phần lớn sinh học phân tử là nghiên cứu định lượng, và trong thời gian gần đây, một lượng lớn công việc đã được thực hiện bằng cách sử dụng các kỹ thuật khoa học máy tính như trong tin sinh học và sinh học tính toán. Di truyền phân tử, hay việc nghiên cứu về cấu trúc và chức năng của gen, là một trong những lĩnh vực phụ nổi bật nhất của sinh học phân tử kể từ đầu những năm 2000. Sinh học phân tử cung cấp thông tin cho các phân ngành sinh học khác theo cả hai cách trực tiếp và gián tiếp. Ở cách trực tiếp, sinh học phân tử nghiên cứu tương tác của các phân tử và những kết quả gặt hái được sau này giúp mở rộng kiến thức cho các phân ngành như sinh học tế bào hay sinh học phát triển. Ở cách gián tiếp, các kỹ thuật trong sinh học phân tử được ứng dụng để tìm các đặc tính lịch sử của các quần thể hoặc loài, và những thông tin có được từ việc làm này giúp phong phú thêm kiến thức trong lĩnh vực sinh học tiến hóa như di truyền học quần thể và phát sinh chủng loài học. Ngoài ra, cũng có một truyền thống lâu đời về việc nghiên cứu các phân tử sinh học ở cấp độ thấp nhất (hay ở cấp độ phân tử) trong phân ngành lý sinh.
Các kỹ thuật trong sinh học phân tử
Truyền tải hình ảnh
Truyền tải hình ảnh là một kỹ thuật được sử dụng để phân lập và chuyển chuỗi ADN mong muốn vào một tác nhân (hay còn gọi là vector) plasmid. Công nghệ ADN tái tổ hợp này đã được phát triển lần đầu vào những năm 1960. Trong kỹ thuật này, trình tự ADN mã hóa cho một protein mong muốn sẽ được nhân bản bằng phản ứng chuỗi polymerase (PCR) và/hoặc các enzym giới hạn để trở thành một plasmid (vector biểu hiện). Vector plasmid thường có ít nhất ba thành phần đặc biệt: điểm bắt đầu sao chép (ori), các vị trí tạo dòng (Multiple Cloning Site hay MCS), và marker chọn lọc (thường là các gen kháng kháng sinh). Ngoài ra, nằm ở vị trí ngược dòng của MCS là vùng gen khởi động và vị trí bắt đầu quá trình phiên mã nhằm điều hòa sự biểu hiện của gen được nhân bản.
Sau khi tổng hợp xong, plasmid có thể được đưa vào tế bào vi khuẩn hoặc động vật. Việc đưa ADN vào tế bào vi khuẩn có thể được thực hiện bằng cách biến nạp thông qua hấp thụ ADN trần, hay tiếp hợp thông qua tiếp xúc giữa tế bào, hoặc tải nạp thông qua vector virus. Việc đưa ADN vào tế bào nhân thực, chẳng hạn như tế bào động vật, bằng các phương pháp vật lý hoặc hóa học được gọi là quá trình chuyển nạp. Có nhiều kỹ thuật chuyển nạp khác nhau như chuyển nạp canxi photphat, điện phân, vi tiêm, hay chuyển nạp liposome. Plasmid có thể được tích hợp vào bộ gen của tế bào chủ và tạo nên hiện tượng chuyển nạp ổn định. Đồng thời, plasmid cũng có thể tồn tại độc lập với bộ gen của tế bào chủ và được biểu hiện tạm thời, và hiện tượng này được gọi là chuyển nạp thoáng qua.
Khi ADN mã hóa cho một protein mong muốn nằm bên trong tế bào, protein đó có thể được biểu hiện. Nhiều yếu tố khác nhau, chẳng hạn như tương tác giữa chất xúc tiến cảm ứng với vùng gen khởi động hay các tín hiệu tế bào cụ thể, giúp quá trình tổng hợp protein mong muốn xảy ra ở một mức độ cao. Sau khi quá trình này hoàn tất, một lượng lớn protein có thể được chiết xuất từ tế bào vi khuẩn hoặc tế bào nhân thực. Protein lúc này có thể được mang đi để kiểm tra hoạt động xúc tác trong nhiều tình huống khác nhau. Ngoài ra, protein có thể được kết tinh để có thể nghiên cứu cấu trúc bậc ba của nó, hoặc, trong ngành dược phẩm, có thể nghiên cứu hoạt tính của các loại thuốc mới đối với protein.
Phản ứng chuỗi polymerase
Phản ứng chuỗi polymerase (PCR) là một kỹ thuật để sao chép DNA có tính ứng dụng cao. Kỹ thuật PCR cho phép sao chép hoặc sửa đổi một trình tự ADN cụ thể theo những cách đã được định sẵn. Phản ứng này cực kỳ công hiệu, và trong điều kiện hoàn hảo có thể khuếch đại một phân tử DNA trở thành 1,07 tỷ phân tử trong vòng chưa đầy hai giờ. PCR có nhiều ứng dụng, bao gồm nghiên cứu biểu hiện gen, phát hiện vi sinh vật gây bệnh, phát hiện đột biến gen, và đưa đột biến vào đoạn DNA. Kỹ thuật PCR có thể được sử dụng để đưa các vị trí enzyme giới hạn vào các đầu của phân tử DNA, hoặc để kích thích đột biến của một base cụ thể trên đoạn DNA thông qua phương pháp gây đột biến có định hướng. PCR cũng có thể được sử dụng để xác định xem một đoạn ADN cụ thể có được tìm thấy trong thư viện ADN bổ sung (cADN) hay không. PCR có nhiều biến thể, như PCR phiên mã ngược (RT-PCR) để khuếch đại ARN, và gần đây là PCR định lượng cho phép đo lường chính xác số phân tử ADN hoặc ARN.

Điện di trên gel
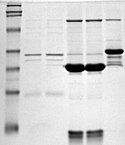
Điện di trên gel là một phương pháp để tách các phân tử dựa trên kích thước của chúng, sử dụng gel agarose hoặc polyacrylamide. Kỹ thuật này là một công cụ quan trọng trong sinh học phân tử. Nguyên lý cơ bản là các đoạn ADN có thể được tách ra dựa trên kích thước khi dòng điện chạy qua gel. Do khung xương ADN chứa các nhóm phosphate mang điện tích âm, ADN sẽ di chuyển về phía dương của dòng điện qua gel agarose. Protein cũng có thể được phân tách theo kích thước bằng gel SDS-PAGE, hoặc theo kích thước và điện tích bằng phương pháp điện di trên gel 2D.
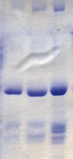
Phương pháp xét nghiệm Bradford
Thử nghiệm Bradford là một kỹ thuật sinh học phân tử cho phép định lượng chính xác và nhanh chóng các phân tử protein, sử dụng thuốc nhuộm đặc biệt là Coomassie Brilliant Blue G-250. Coomassie Blue thay đổi màu từ nâu đỏ sang xanh lam khi liên kết với protein. Protein kết nối với Coomassie Blue trong môi trường axit, làm thay đổi bước sóng từ 465 nm sang 595 nm, tạo ra màu xanh lam sáng. Sau khi phản ứng, nồng độ protein trong thử nghiệm Bradford có thể được đo bằng máy quang phổ mà không cần thiết bị đắt tiền.
Phương pháp này được phát triển vào năm 1975 bởi Marion M. Bradford, cho phép định lượng protein nhanh chóng và chính xác hơn nhiều so với các phương pháp trước đó như quy trình Lowry hay xét nghiệm biuret. Không giống như các phương pháp trước đây, xét nghiệm Bradford ít bị ảnh hưởng bởi các chất không phải protein như ethanol, natri clorua hoặc magie clorua. Tuy nhiên, phương pháp này có thể bị ảnh hưởng bởi các dung dịch đệm mạnh như natri dodecyl sulfat (SDS).
Các phương pháp thấm và phương pháp sàng lọc các đại phân tử
Các thuật ngữ Northern (phương bắc), Western (phương tây) và Eastern (phương đông) ban đầu là từ một trò đùa trong ngành sinh học phân tử khi các nhà khoa học đặt tên để đùa với thuật ngữ Southern blotting (phương pháp thấm Southern), theo Edwin Southern mô tả để lai ghép ADN. Patricia Thomas, nhà phát triển phương pháp thấm ARN sau này gọi là Northern blot (phương pháp thấm Northern), thực tế không sử dụng thuật ngữ này.
Phương pháp thấm Southern
Phương pháp thấm Southern được đặt theo tên nhà sinh học Edwin Southern, người phát minh ra kỹ thuật này. Đây là phương pháp để thăm dò sự có mặt của một đoạn ADN cụ thể trong một mẫu ADN hỗn hợp. ADN mẫu sau khi cắt bằng enzym giới hạn sẽ được tách bằng phương pháp điện di trên gel và chuyển sang màng lọc bằng thẩm mao dẫn. Màng lọc này sau đó tiếp xúc với một đầu dò ADN được đánh dấu chứa trình tự gen có thể liên kết với đoạn ADN quan tâm. Phương pháp thấm Southern hiện nay ít được sử dụng hơn trong phòng thí nghiệm khoa học do sự ưu việt của các kỹ thuật khác như PCR để phát hiện trình tự ADN cụ thể từ các mẫu ADN. Tuy nhiên, phương pháp này vẫn được áp dụng trong một số ứng dụng như đo lượng bản sao gen trong chuột biến đổi gen hoặc trong kỹ thuật tạo dòng tế bào gốc phôi thông qua gen knockout.
Phương pháp thấm Northern
Phương pháp thấm Northern được áp dụng để nghiên cứu sự hiện diện của các phân tử ARN cụ thể để so sánh tương đối giữa các mẫu ARN khác nhau. Cơ bản, kỹ thuật này kết hợp giữa phương pháp điện di của ARN trên gel với phương pháp thấm. Trong quá trình này, ARN được phân tách theo kích thước và sau đó chuyển đến màng lọc trước khi thăm dò với một trình tự quan tâm được đánh dấu. Kết quả có thể được hình dung thông qua nhiều phương pháp khác nhau dựa vào chất đánh dấu sử dụng. Tuy nhiên, thường nhìn thấy các dải biểu thị kích thước của ARN được phát hiện trong mẫu. Cường độ của các dải này liên quan đến lượng ARN mục tiêu trong các mẫu được phân tích. Quy trình này thường được áp dụng để nghiên cứu thời điểm và mức độ biểu hiện gen bằng cách đo lượng ARN hiện diện trong các mẫu khác nhau, giả sử rằng không có quá trình điều hòa sau khi phiên mã và mức độ ARN phản ánh tỷ lệ sản xuất protein tương ứng.
Phương pháp thấm Western
Phương pháp thấm Western là kỹ thuật cho phép phát hiện các protein cụ thể từ một hỗn hợp các protein. Kỹ thuật này cũng cho phép xác định kích thước của các protein được tách riêng và định lượng sự biểu hiện của chúng. Trong phương pháp thấm Western, protein được tách ra theo kích thước trên một gel SDS-PAGE và sau đó chuyển sang màng PVDF, nitrocellulose, nylon hoặc các màng lọc khác. Màng lọc này sau đó có thể được thăm dò bằng các dung dịch kháng thể. Các kháng thể liên kết với protein quan tâm có thể được hình dung thông qua nhiều kỹ thuật khác nhau như sử dụng các phản ứng màu, phản ứng phát quang hóa học hoặc chất phóng xạ tự chụp. Thông thường, các kháng thể được đánh dấu bằng enzym. Khi tiếp xúc với chất nền phát quang hóa học, tổ hợp này có thể được phát hiện. Sử dụng phương pháp thấm Western không chỉ cho phép phát hiện mà còn cho phép phân tích định lượng. Các phương pháp tương tự có thể được sử dụng để nhuộm trực tiếp các protein cụ thể trong tế bào sống hoặc các phần mô.
Phương pháp thấm Eastern
Phương pháp thấm Eastern được áp dụng để phát hiện các biến đổi sau quá trình dịch mã của protein. Trong kỹ thuật này, protein thấm trên màng PVDF hoặc nitrocellulose được kết hợp với các chất nền cụ thể để thăm dò các sửa đổi của protein sau khi dịch mã.
Kỹ thuật ADN microarray
Một ADN microarray hoặc ADN chip là một tập hợp các điểm được gắn trên một nền vững chắc như kính hiển vi, trong đó mỗi điểm chứa một hoặc nhiều đoạn oligonucleotide ADN đơn. Các array giúp đặt một số lượng lớn các điểm rất nhỏ (đường kính 100 micromet) trên một mặt kính duy nhất. Mỗi điểm chứa một phân tử đoạn ADN phụ thuộc vào một chuỗi ADN đơn. Một biến thể của kỹ thuật này cho phép biểu hiện gen của một sinh vật ở một giai đoạn phát triển cụ thể. Trong kỹ thuật này, ARN trong mô được phân lập và chuyển đổi thành ADN phụ thuộc (cADN) được đánh dấu. cADN này sau đó được lai với các đoạn trên mảng, và hình ảnh của quá trình lai có thể được ghi lại. Vì nhiều dãy có thể được tạo thành từ các đoạn gen giống nhau, điều này rất hữu ích để so sánh biểu hiện gen của hai mô khác nhau như mô khỏe mạnh và mô ung thư. Ngoài ra, có thể đo lường sự biểu hiện gen và biểu hiện thay đổi theo thời gian hoặc các yếu tố khác. Có nhiều cách để chế tạo các dãy, phổ biến nhất là các chip silicon, lam kính với các điểm có đường kính ~100 micromet, các dãy được tinh chỉnh và các dãy với các điểm lớn trên bề mặt một tấm màng với nhiều lỗ (macroarrays). Có từ 100 điểm đến hơn 10,000 điểm trên một dãy nhất định. Dãy cũng có thể được tạo bằng các phân tử khác ngoài ADN.
Oligonucleotide đặc hiệu alen
Oligonucleotide đặc hiệu alen (ASO) là một kỹ thuật cho phép phát hiện các đột biến cơ sở đơn mà không cần dùng đến PCR hoặc điện di trên gel. Các mẫu dò ngắn (chiều dài khoảng 20-25 nucleotide) được đánh dấu và tiếp xúc với ADN mục tiêu không phân mảnh. Quá trình lai xảy ra với độ đặc hiệu cao do độ dài ngắn của các dây dò, và thậm chí một thay đổi của một base đơn lẻ cũng có thể ảnh hưởng đến quá trình lai. ADN mục tiêu sau đó được rửa sạch để loại bỏ các dây dò đã đánh dấu nhưng không lai. ADN mục tiêu sau đó được phân tích để xác định sự hiện diện của các dây dò thông qua phóng xạ hoặc huỳnh quang. Như trong hầu hết các kỹ thuật sinh học phân tử khác, một nhóm kiểm soát phải được sử dụng để đảm bảo thành công của thí nghiệm.
Trong lĩnh vực sinh học phân tử, các quy trình và công nghệ liên tục được phát triển và các công nghệ cũ đang bị loại bỏ. Ví dụ, trước khi kỹ thuật điện di trên gel (agarose hoặc polyacrylamide) cho ADN được phát triển, kích thước của các phân tử ADN thường được xác định bằng cách đo tốc độ lắng trong gradient đường sucroza, một kỹ thuật chậm chạp, tốn nhiều công sức và sử dụng các thiết bị đo đắt tiền. Trước đó, máy đo độ nhớt đã được sử dụng để đo kích thước phân tử ADN. Đôi khi, việc hiểu biết về các công nghệ cũ là cần thiết vì những kỹ thuật này có thể hữu ích trong việc giải quyết các vấn đề mới mà các công nghệ tiên tiến không thể làm được.