



Sự kiện tuyệt chủng Phấn Trắng – Kỷ Cổ Cận (K–Pg), còn được gọi là sự kiện tuyệt chủng Creta–Paleogen hoặc sự kiện tuyệt chủng Phấn Trắng – Đệ Tam (K–T), là một đại tuyệt chủng xảy ra cách đây khoảng 66 triệu năm vào cuối thời kỳ Maastricht, khi hàng loạt loài động thực vật bị tuyệt chủng trong một khoảng thời gian địa chất ngắn. Sự kiện này còn liên quan đến ranh giới địa chất giữa kỷ Phấn Trắng và kỷ Cổ Cận, đó là một lớp trầm tích mỏng có mặt ở nhiều khu vực trên thế giới. Sự kiện này đánh dấu sự kết thúc của đại Trung Sinh và sự khởi đầu của đại Tân Sinh.
Hóa thạch của các loài khủng long không phải chim chỉ xuất hiện dưới ranh giới K-T, cho thấy rằng tất cả các loài khủng long không thuộc về nhóm chim đã tuyệt chủng trong sự kiện này. Một số ít hóa thạch khủng long xuất hiện trên ranh giới K-T, nhưng điều này có thể do quá trình tái lắng đọng, tức là hóa thạch bị xói mòn từ nơi nguyên thủy và sau đó được bảo tồn trong các lớp trầm tích mới hình thành. Các loài như thương long, thằn lằn cổ rắn, thằn lằn có cánh, cùng một số thực vật và động vật không xương sống cũng biến mất. Nhánh động vật có vú sống sót qua sự kiện này với số lượng nhỏ bị tuyệt chủng, và sự phân tách tiến hóa từ các nhánh có mặt trong tầng Maastricht phát triển mạnh mẽ sau ranh giới này. Tốc độ tuyệt chủng và phân nhánh thay đổi giữa các nhóm sinh vật.
Các nhà khoa học đưa ra giả thuyết rằng sự kiện tuyệt chủng K–T có thể do một hoặc nhiều thảm họa gây ra, như sự va chạm mạnh mẽ của các thiên thạch (như hố Chicxulub) hoặc do hoạt động núi lửa gia tăng. Một số hố va chạm và hoạt động núi lửa đã được xác định tuổi tương ứng với thời điểm sự kiện tuyệt chủng. Những sự kiện địa chất này có thể làm giảm ánh sáng mặt trời và mức độ quang hợp, dẫn đến sự phá hủy hệ sinh thái trên quy mô lớn. Một số nghiên cứu khác cho rằng sự tuyệt chủng diễn ra từ từ, do sự thay đổi chậm chạp của mực nước biển hoặc khí hậu.
Những kiểu tuyệt chủng
Mặc dù sự kiện này đánh dấu một ranh giới địa chất khốc liệt, nhưng nó cũng thể hiện sự thay đổi đáng kể về tốc độ tuyệt chủng giữa và trong các nhóm sinh vật khác nhau. Các loài phụ thuộc vào quang hợp bị suy giảm hoặc tuyệt chủng do các hạt vật chất trong khí quyển che khuất ánh sáng mặt trời và làm giảm năng lượng mặt trời đến bề mặt Trái Đất. Sự tuyệt chủng của thực vật gây ra sự xáo trộn lớn trong các nhóm thực vật chi phối. Các loài sinh vật quang hợp, bao gồm thực vật phiêu sinh và thực vật trên cạn, đã là nền tảng của chuỗi thức ăn vào cuối kỷ Phấn Trắng, giống như ngày nay. Các dấu vết cho thấy các động vật ăn cỏ đã chết khi nguồn thực phẩm chính của chúng trở nên khan hiếm, dẫn đến cái chết của các động vật săn mồi hàng đầu như Tyrannosaurus rex.
Tảo gai vôi (Coccolithophore) và động vật thân mềm (bao gồm cúc đá, Rudistes, ốc và trai nước ngọt, cùng các sinh vật ăn tảo) bị tuyệt chủng hoặc chịu tổn thất nghiêm trọng. Ví dụ, cúc đá được cho là thức ăn chính của thương long (Mosasauridae), một nhóm bò sát biển khổng lồ đã biến mất trong sự kiện này.
Các loài động vật ăn tạp, động vật ăn sâu bọ và động vật ăn xác chết đã sống sót qua sự kiện tuyệt chủng, có thể nhờ vào sự gia tăng nguồn thực phẩm. Cuối kỷ Phấn Trắng, dường như không còn các loài động vật có vú ăn cỏ hay ăn thịt thuần túy. Thú và chim sống sót đã ăn côn trùng, giun và ốc, những sinh vật này lại tiêu thụ động vật và thực vật chết. Các nhà khoa học cho rằng những sinh vật này đã sống sót sau sự sụp đổ của chuỗi thức ăn dựa trên thực vật nhờ việc tiêu thụ thức ăn vụn (vật chất hữu cơ không còn sống).
Trong các quần xã sinh vật ở suối, chỉ một số nhóm động vật bị tuyệt chủng do sự phụ thuộc ít vào thực vật sống và nhiều vào thức ăn mảnh vụn từ đất liền, giúp chúng tránh khỏi sự tuyệt chủng. Các mô hình tương tự, nhưng phức tạp hơn, xuất hiện trong các đại dương. Sự tuyệt chủng nghiêm trọng hơn đối với động vật sống ở tầng nước mặt so với những động vật ở đáy biển. Động vật trong cột nước chủ yếu phụ thuộc vào thực phẩm từ thực vật trôi nổi, trong khi động vật sống ở đáy đại dương ăn mảnh vụn hữu cơ hoặc chuyển sang ăn mảnh vụn.
Các loài động vật thở không khí lớn nhất sống sót qua sự kiện này, như cá sấu (Crocodyliformes) và li long (Choristodera), là những dạng bán thủy sinh có thể tiếp cận nguồn thức ăn mảnh vụn. Cá sấu hiện đại có khả năng sống như động vật ăn xác chết và có thể tồn tại vài tháng mà không cần thức ăn, trong khi con non của chúng thì nhỏ, phát triển chậm và chủ yếu ăn động vật không xương sống, sinh vật chết hoặc mảnh vụn trong những năm đầu đời. Những đặc điểm này có thể giải thích cho sự sống sót của cá sấu vào cuối kỷ Phấn Trắng.
Sau sự kiện K–T, sự đa dạng sinh học cần thời gian dài để phục hồi, mặc dù có rất nhiều hốc sinh thái bị bỏ trống.
Vi sinh vật
Ranh giới K-T biểu thị một trong những sự thay đổi lớn nhất trong hóa thạch của các vi phiêu sinh vật chứa vôi, hình thành nên trầm tích calci, và là nguồn gốc của tên gọi kỷ Phấn trắng (kỷ Phấn Trắng). Sự chuyển đổi trong nhóm này rất rõ ràng ở cấp độ loài. Phân tích thống kê về sự suy giảm sinh vật biển trong thời kỳ này cho thấy sự giảm đa dạng sinh học chủ yếu là do sự gia tăng mạnh mẽ của tuyệt chủng, hơn là do sự giảm hình thành loài. Dấu vết về giáp tảo (Dinoflagellata) trong ranh giới K-T vẫn chưa được hiểu rõ, vì chỉ có các bào xác vi sinh vật được ghi nhận trong hóa thạch, và không phải tất cả các loài giáp tảo đều có giai đoạn tạo bào xác, do đó sự đa dạng có thể đã bị đánh giá thấp. Các nghiên cứu gần đây cho thấy không có sự thay đổi lớn nào đối với giáp tảo trong lớp trầm tích của ranh giới này.
Trùng tia đã xuất hiện trong các lớp địa chất ít nhất từ kỷ Ordovic, và các khung xương hóa thạch khoáng hóa của chúng có thể được thấy trong ranh giới K–T. Không có dấu hiệu của sự tuyệt chủng lớn của các loài này, và có bằng chứng cho thấy sự sinh sôi nảy nở mạnh mẽ ở các loài này tại các vĩ độ cao ở bán cầu nam khi nhiệt độ giảm dần vào Paleocen sớm. Khoảng 46% các loài tảo cát đã sống sót qua giai đoạn này từ Creta đến Paleocen thượng. Điều này cho thấy có sự luân chuyển đáng kể về loài, thay vì sự tuyệt chủng thảm họa của các loài tảo cát trong suốt ranh giới K–T.
Sự xuất hiện của phiêu sinh vật trùng lỗ trong ranh giới K–T đã được nghiên cứu từ thập niên 1930. Nghiên cứu này được thúc đẩy bởi khả năng có một sự kiện gây ảnh hưởng tại ranh giới K–T đã dẫn đến một loạt các ấn phẩm miêu tả chi tiết sự tuyệt chủng của phiêu sinh trùng lỗ ở ranh giới này. Tuy nhiên, vấn đề này vẫn còn gây tranh cãi giữa các nhà khoa học, với một nhóm cho rằng có dấu hiệu của sự tuyệt chủng đáng kể về số lượng loài này ở ranh giới K–T, trong khi nhóm khác tin rằng có nhiều lần tuyệt chủng và nở rộ của các loài này tại ranh giới K-T.
Nhiều loài trùng lỗ sống tầng đáy đã bị tuyệt chủng trong sự kiện K–Pg, theo kết luận dựa trên các mảnh vụn hữu cơ từ chất dinh dưỡng do sinh khối đại dương suy giảm. Tuy nhiên, khi các vi sinh vật biển hồi phục, người ta cho rằng sự phân hóa các loài trùng lỗ gia tăng là do nguồn thức ăn tăng lên. Sự hồi phục của phiêu thực vật vào Paleocen sớm đã cung cấp nguồn thức ăn cho các chủng loài trùng lỗ tầng đáy, những loài ăn mảnh vụn. Sự phục hồi cuối cùng về số lượng các loài sống đáy đã diễn ra qua nhiều giai đoạn kéo dài hàng trăm ngàn năm đến Paleocen sớm.
Động vật không xương sống ở biển

Sự biến động trong hồ sơ hóa thạch và tốc độ tuyệt chủng của các loài động vật không xương sống biển trong thời kỳ ranh giới K-T đã xảy ra. Tốc độ biểu kiến bị ảnh hưởng nhiều hơn bởi sự thiếu vắng hồ sơ hóa thạch hơn là sự tuyệt chủng thực sự của các loài.
Bọ nước (Ostracoda), một nhóm nhỏ giáp xác phổ biến trong tầng Maastricht thượng, đã để lại trầm tích hóa thạch ở nhiều khu vực khác nhau. Phân tích các hóa thạch này cho thấy sự đa dạng của Ostracoda trong thế Paleocen thấp hơn so với các thời kỳ khác trong kỷ đệ Tam. Tuy nhiên, hiện tại chưa rõ liệu sự tuyệt chủng đã xảy ra trước hay trong quá trình hình thành ranh giới này.
Khoảng 60% các chi san hô đá (Scleractinia) từ kỷ Creta muộn đã không vượt qua được ranh giới K–T để bước sang thế Paleocen. Phân tích cho thấy khoảng 98% các loài san hô tập đoàn, sống trong môi trường nước ấm, nông, cận nhiệt đới, đã bị tuyệt chủng. Ngược lại, các loài san hô đơn độc, sống trong môi trường nước lạnh và sâu hơn (dưới đới sáng rõ) ít bị ảnh hưởng hơn tại ranh giới K–T. Các loài san hô tập đoàn phụ thuộc vào tảo quang hợp, và sự kiện này đã dẫn đến cái chết của các loài tảo đó. Tuy nhiên, việc sử dụng dữ liệu từ các hóa thạch san hô để giải thích sự kiện tuyệt chủng K-T và sự phục hồi sau đó trong thế Paleocen cần cân nhắc với những thay đổi trong hệ sinh thái san hô xuyên suốt ranh giới K–T.
Số lượng các chi động vật chân đầu, động vật da gai, và thân mềm hai mảnh vỏ đã giảm đáng kể sau ranh giới K-T. Hầu hết các loài động vật tay cuộn (Brachiopoda), một nhóm nhỏ của động vật không xương sống biển, đã sống sót qua sự kiện K–T và tiếp tục đa dạng hóa trong thời kỳ Paleocen sớm.

Ngoại trừ các loài nautiloidea (với đại diện hiện đại là bộ nautilida) và coleoidea (tách thành Octopoda, Teuthida, và Sepiida hiện đại), tất cả các loài động vật thâm mềm khác thuộc lớp Chân đầu đều bị tuyệt chủng tại ranh giới K-T. Các loài như tiễn thạch (Belemnoidea) và cúc đá, nhóm động vật chân đầu có vỏ với sự đa dạng cao về số lượng và phân bố rộng rãi, cũng không còn tồn tại. Các nghiên cứu cho thấy nautiloidea còn sống sót nhờ vào số lượng trứng ít nhưng lớn, điều này giúp chúng duy trì sự sống còn qua sự kiện tuyệt chủng, trong khi cúc đá với cách sinh sản dạng phiêu sinh (nhiều trứng và ấu trùng phiêu sinh) có thể đã bị hủy diệt trong sự kiện này. Nghiên cứu thêm cho thấy sau khi Ammonoidea bị loại khỏi hệ sinh thái toàn cầu, Nautiloidea đã tiến hóa thành các dạng phức tạp hơn với vỏ, dù đặc điểm này chỉ thấy ở cúc đá.
Khoảng 35% các chi động vật da gai đã bị tuyệt chủng tại ranh giới K-T. Mặc dù các loài da gai trước đây phát triển mạnh trong môi trường nước nông, vĩ độ thấp trong thời kỳ Creta muộn, tỷ lệ tuyệt chủng của chúng lại cao. Ngược lại, các loài da gai sống ở môi trường nước sâu và vĩ độ trung bình ít bị ảnh hưởng hơn. Sự tuyệt chủng chủ yếu là do mất môi trường sống, đặc biệt là sự phá hủy các rạn san hô nước nông do sự kiện tuyệt chủng.
Các nhóm động vật không xương sống khác như rudist (nghêu tạo rạn san hô) và Inoceramus (loài khổng lồ liên quan đến sò (Pectinidae) hiện đại) cũng đã bị tuyệt chủng tại ranh giới K-T.
Cá
Trong ranh giới K-T, có một số lượng đáng kể các hóa thạch cá có quai hàm, cho thấy kiểu tuyệt chủng của các lớp động vật có xương sống. Trong nhóm cá sụn, khoảng 80% các họ cá mập và cá đuối đã sống sót qua sự kiện này, và hơn 90% các họ cá xương (teleostei) cũng sống sót. Có bằng chứng cho thấy số lượng lớn hóa thạch cá xương được bảo tồn tại các địa điểm hóa thạch nằm ngay trên lớp ranh giới K-T ở đảo Seymour gần Nam Cực, dường như phản ánh sự lắng đọng từ sự kiện K–Pg. Tuy nhiên, các loài cá sống trong môi trường nước ngọt và nước mặn chịu ít tác động hơn từ sự kiện tuyệt chủng.
Động vật không xương sống trên cạn
Tổn thương do côn trùng gây ra trên lá hóa thạch của thực vật có hoa tại 14 di chỉ ở Bắc Mỹ được dùng để đại diện cho sự đa dạng của côn trùng ở ranh giới K-T và để phân tích tốc độ tuyệt chủng. Các nhà nghiên cứu phát hiện rằng ở các di chỉ có niên đại từ kỷ Creta, trước sự kiện tuyệt chủng, đa dạng của các loài ăn thực vật và côn trùng rất phong phú. Tuy nhiên, trong thời kỳ Paleocen sớm, hệ thực vật vẫn còn đa dạng nhưng chỉ có một số ít loài ăn côn trùng, tình trạng này kéo dài tới 1,7 triệu năm sau sự kiện tuyệt chủng.
Thực vật trên cạn
Có bằng chứng rõ ràng về sự tàn phá các quần thể thực vật tại ranh giới K-T. Tuy nhiên, mức độ tàn phá và sự diễn thế thực vật khác nhau giữa các khu vực. Ở Bắc Mỹ, dữ liệu cho thấy sự phá hủy đáng kể hệ thực vật và sự tuyệt chủng lớn của thực vật trong các phần của ranh giới K-T, mặc dù đã có những thay đổi lớn trong các hệ thực vật trước ranh giới này.
Tại các vĩ độ cao của Nam bán cầu như New Zealand và Nam Cực, hệ thực vật bị tàn phá nghiêm trọng, dẫn đến sự hồi phục không đáng kể của một số loài, mặc dù có sự thay đổi ngắn hạn trong sự phong phú của các nhóm thực vật. Ở Bắc Mỹ, khoảng 57% các loài thực vật đã tuyệt chủng. Trong thời kỳ Paleocen, sự hồi phục thực vật bắt đầu với sự xâm nhập của dương xỉ, được thể hiện qua đỉnh dương xỉ trong các lớp địa tầng, tương tự như sự tái chiếm của dương xỉ sau vụ phun trào núi St. Helen năm 1980.
Với sự tàn phá tổng thể của các loài thực vật ở ranh giới K-T, nấm, các sinh vật hoại sinh không cần quang hợp và sử dụng chất dinh dưỡng từ thực vật phân hủy, đã gia tăng. Nấm thống trị chỉ trong vài năm khi bầu khí quyển được làm sạch và có nhiều chất hữu cơ để tiêu thụ. Khi bầu khí quyển đã trở lại bình thường, các sinh vật quang hợp như dương xỉ và các loài thực vật khác bắt đầu hồi sinh. Tính đa bội dường như đã giúp thực vật có hoa sống sót qua sự tuyệt chủng, nhờ các bản sao gen bổ sung cho phép chúng dễ dàng thích nghi với các điều kiện môi trường thay đổi nhanh chóng sau sự va chạm.
Động vật lưỡng cư
Không có dấu hiệu cho thấy các loài lưỡng cư bị tuyệt chủng lớn ở ranh giới K-T, và có bằng chứng mạnh mẽ rằng phần lớn các loài lưỡng cư đã sống sót tương đối nguyên vẹn sau sự kiện này. Nghiên cứu chi tiết về các chi kỳ giông trong các lớp hóa thạch ở Montana cho thấy 6 trong số 7 chi không có sự thay đổi đáng kể sau sự kiện.
Các loài ếch nhái gần như sống sót đến Paleocen với chỉ một số ít loài tuyệt chủng. Tuy nhiên, hồ sơ hóa thạch về các họ và chi ếch nhái là không đồng đều. Nghiên cứu mở rộng về 3 chi ếch ở Montana cho thấy không có ảnh hưởng rõ rệt từ sự kiện K–Pg và sự sống sót dường như không thay đổi. Dữ liệu chỉ ra ít hoặc không có dấu hiệu về sự tuyệt chủng của các loài lưỡng cư liên quan đến sự kiện K–Pg. Sự sống sót của lưỡng cư có thể là do khả năng tự tìm nơi trú ẩn trong nước hoặc xây dựng hang hốc trong trầm tích, đất, gỗ hoặc dưới các tảng đá.
Bò sát không thuộc Archosauria

Hai nhóm bò sát không thuộc nhánh Archosauria (thằn lằn chúa), gồm Testudines (rùa) và Lepidosauria (thằn lằn vảy, bao gồm rắn, thằn lằn, và thằn lằn giun), cùng với ly long (Choristodera, nhóm bò sát dạng thằn lằn chúa (Archosauromorpha) sống bán thủy sinh, đã tuyệt chủng vào Miocen sớm), đã sống sót qua ranh giới K-T. Hơn 80% các loài rùa từ kỷ Phấn Trắng đã sống sót qua sự kiện này. Tất cả sáu họ rùa hiện diện vào cuối kỷ Phấn Trắng đều tồn tại đến kỷ đệ Tam và trở thành các loài rùa hiện đại.
Các loài thằn lằn vảy hiện còn bao gồm Rhynchocephalia (tuatara) và Squamata (bò sát có vảy). Rhynchocephalia từng là nhóm thằn lằn vảy phổ biến vào đầu đại Trung Sinh nhưng suy giảm từ giữa kỷ Phấn Trắng. Hiện tại, chúng chỉ còn một chi duy nhất ở New Zealand.
Nhóm Squamata, bao gồm thằn lằn, rắn và thằn lằn giun, đã phân nhánh vào các hốc sinh thái khác nhau trong suốt kỷ Jura và phát triển mạnh mẽ suốt kỷ Phấn Trắng. Chúng đã sống sót qua ranh giới K-T và hiện là nhóm đa dạng nhất trong số các loài bò sát với hơn 6.000 loài. Không có họ bò sát có vảy trên cạn nào đã tuyệt chủng tại ranh giới này, và dấu hiệu hóa thạch cho thấy chúng không trải qua sự suy giảm lớn về số lượng. Kích thước nhỏ, khả năng thích nghi trao đổi chất, và khả năng di chuyển đến các môi trường phù hợp hơn là những yếu tố quan trọng giúp chúng tồn tại qua kỷ Creta muộn và Paleogen sớm.
Các loài bò sát khổng lồ sống dưới nước không thuộc nhóm thằn lằn chúa, chẳng hạn như Mosasauroidea và Plesiosauria, là những kẻ săn mồi hàng đầu trong đại dương thời kỳ đó, đã biến mất vào cuối kỷ Phấn Trắng.
Bò sát Archosauria (Thằn lằn chúa)
Nhánh thằn lằn chúa (Archosauria) bao gồm hai bộ hiện còn tồn tại: cá sấu (với các họ hiện còn là Alligatoridae, Crocodylidae và Gavialidae) và khủng long (Dinosauria, trong đó chim là nhóm duy nhất còn sống sót), cùng với các loài khủng long bay và thằn lằn có cánh (Pterosauria) đã tuyệt chủng.
Bò sát dạng cá sấu
Có 10 họ bò sát dạng cá sấu (Crocodiliformes) hoặc họ hàng gần của chúng được tìm thấy trong các hóa thạch của tầng Maastricht, trong đó 5 họ đã tuyệt chủng trước ranh giới K-T. 5 họ còn lại có hóa thạch trong các trầm tích Maastricht và Paleocen. Các họ còn sống sót của Crocodyliformes hiện sống ở môi trường nước ngọt và trên cạn, ngoại trừ Dyrosauridae sống ở cả nước ngọt và nước mặn. Khoảng 50% đại diện của Crocodyliformes đã sống sót qua ranh giới K-T, với xu hướng rõ ràng là không có cá sấu lớn sống sót. Sự sống sót của bò sát dạng cá sấu qua ranh giới K-T có thể do khả năng thích nghi của chúng với môi trường nước và đào hang, giảm thiểu sự nhạy cảm với tác động môi trường tiêu cực trong sự kiện này. Vào năm 2008, Jouve và cộng sự đã gợi ý rằng các loài bò sát dạng cá sấu nhỏ sống trong môi trường nước ngọt như các loài cá sấu biển hiện đại đã giúp chúng sống sót khi các loài bò sát biển khác tuyệt chủng, vì môi trường nước ngọt ít bị ảnh hưởng trong sự kiện K–Pg.
Thằn lằn có cánh
Chỉ có một họ thằn lằn có cánh (Pterosauria), cụ thể là Azhdarchidae, được ghi nhận rõ ràng trong tầng Maastricht, và đã tuyệt chủng tại ranh giới K-T. Những thằn lằn có cánh lớn này là các đại diện cuối cùng của nhóm đã từng gồm khoảng 10 họ giữa kỷ Phấn Trắng. Các loài thằn lằn có cánh nhỏ hơn đã tuyệt chủng trước tầng Maastricht trong thời gian nhóm động vật nhỏ suy giảm và động vật lớn trở nên phổ biến hơn. Trong giai đoạn này, các loài chim hiện đại phát triển đa dạng và thay thế các nhóm chim cổ và thằn lằn có cánh, có thể do cạnh tranh trực tiếp hoặc đơn giản là lấp đầy các hốc sinh thái trống.
Chim
Hầu hết các nhà cổ sinh vật học cho rằng chim là nhánh duy nhất còn tồn tại của khủng long (xem bài nguồn gốc của chim). Tuy nhiên, tất cả các loài chim không thuộc nhóm chim hiện đại (Neornithes) đã tuyệt chủng, bao gồm các nhóm thịnh vượng như Enantiornithes (chim đối ngược) và Hesperornithes (chim hoàng hôn). Một số phân tích hóa thạch chim cho thấy có sự phân nhánh trước ranh giới K-T và cho rằng các họ hàng của vịt, gà và đà điểu đã sống cùng với các loài khủng long phi chim. Một số lượng lớn mẫu hóa thạch chim cho thấy sự tồn tại của các loài chim cổ trong khoảng 300.000 năm của ranh giới K–Pg. Không có loài nào trong số đó được biết đã sống đến thế Paleogen, do đó chứng minh sự tuyệt chủng hàng loạt của chim cổ trùng khớp với sự va chạm của tiểu hành tinh Chicxulub. Một số loài chim kỷ Phấn Trắng đã sống sót qua sự kiện này và phát triển thành các loài chim hiện nay. Hơn nữa, chỉ một loài chim chưa được đặt tên đã chắc chắn được xác định tồn tại trên và dưới ranh giới K-Pg (trong hệ tầng Hell Creek thuộc tầng Maastricht và hệ tầng Fort Union thuộc tầng Đan Mạch).
Nhóm chim duy nhất có thể khẳng định đã sống sót qua ranh giới K-T là chim hiện đại (Neornithine), ngoại trừ một loài trong thế Paleogen, Qinornis paleocenica, có thể không thuộc nhóm Neornithine. Chim hiện đại có khả năng tồn tại sau sự kiện tuyệt chủng nhờ vào khả năng lặn, bơi, và tìm nơi cư trú trong nước và đầm lầy. Một số loài chim hiện đại còn có thể đào hang hoặc làm tổ trong hốc cây và tổ mối, giúp chúng ít bị ảnh hưởng bởi biến đổi môi trường trong thời kỳ này. Sự sống sót kéo dài qua ranh giới này chủ yếu là do việc lấp đầy các hốc sinh thái trống sau sự tuyệt chủng của khủng long phi điểu.
Khủng long phi điểu
Hầu hết các nhà khoa học đồng ý rằng tất cả các loài khủng long phi điểu đã tuyệt chủng ở ranh giới K-T, mặc dù còn một số tranh cãi. Các hóa thạch khủng long cho thấy sự sụt giảm đa dạng hoặc không sụt giảm trong vài triệu năm cuối của kỷ Phấn Trắng, có thể do chất lượng hóa thạch không đủ tốt. Vì không có bằng chứng cho thấy khủng long phi điểu trong tầng Maastricht muộn có khả năng đào hang, bơi hoặc lặn, nên chúng không thể bảo vệ mình khỏi những tác động môi trường nghiêm trọng xảy ra ở ranh giới K-T. Có thể các loài khủng long nhỏ đã sống sót, nhưng chúng có thể thiếu thức ăn, do nhóm khủng long ăn cỏ thiếu thực vật, còn khủng long ăn thịt gặp khó khăn với nguồn cung con mồi.
Sự đồng thuận về nội nhiệt của khủng long giúp giải thích sự tuyệt chủng hoàn toàn của chúng khi so sánh với các họ hàng gần gũi như cá sấu. Các loài cá sấu ngoại nhiệt ('máu lạnh') cần ít thức ăn hơn và có thể sống lâu mà không ăn, trong khi các loài nội nhiệt ('máu nóng') với kích thước tương tự cần nhiều thức ăn hơn để duy trì sự trao đổi chất nhanh chóng. Khi chuỗi thức ăn bị phá vỡ, khủng long phi điểu bị tiêu diệt trong khi một số cá sấu sống sót. Sự sống sót của các loài nội nhiệt khác như thú và chim có thể là do nhu cầu thức ăn ít hơn hoặc kích thước nhỏ hơn tại thời điểm tuyệt chủng.
Việc tuyệt chủng xảy ra từ từ hay đột ngột vẫn là một chủ đề gây tranh cãi, vì cả hai quan điểm đều có bằng chứng hóa thạch hỗ trợ. Một nghiên cứu năm 2010 về 29 địa điểm hóa thạch ở dãy núi Pyrenees thuộc Catalunya, Tây Ban Nha, cho rằng khủng long đã phát triển rất đa dạng cho đến thời điểm thiên thạch va chạm vào Trái Đất. Ngược lại, các hóa thạch ở dọc sông Red Deer, Alberta, Canada, cho thấy sự giảm dần số lượng loài khủng long phi điểu từ khoảng 45 xuống còn 12 trong 10 triệu năm cuối của kỷ Creta, hỗ trợ quan điểm về sự tuyệt chủng từ từ. Một số nhà khoa học khác cũng đưa ra quan điểm tương tự.

Một số nhà nghiên cứu bảo vệ quan điểm khủng long sống sót qua kỷ Paleocen dựa vào việc phát hiện các dấu tích khủng long trong hệ tầng Hell Creek dày 1,3 m, nằm trên và trẻ hơn 40.000 năm so với ranh giới K-T. Các mẫu phấn hoa thu thập gần xương đùi khủng long mỏ vịt (Hadrosauridae) trong sa thạch Ojo Alamo Sandstone ở sông San Juan cho thấy loài này sống vào kỷ Đệ Tam, khoảng 64,5 triệu năm trước (khoảng 1 triệu năm sau sự kiện K-Pg). Nếu sự tồn tại này qua ranh giới K-T được xác nhận, các loài khủng long mỏ vịt có thể được xem là Dead Clade Walking. Một nghiên cứu hiện tại cũng cho thấy các hóa thạch này bị xói mòn từ vị trí nguyên thủy và sau đó được tái lắng đọng trong các lớp trầm tích trẻ hơn.
Thú có vú
Tất cả các nhánh thú kỷ Phấn Trắng như Monotremata (thú đẻ trứng), Multituberculata (thú răng nhiều mấu), Marsupialia (thú có túi) và Eutheria (thú thật sự), cùng với Dryolestoidea và Gondwanatheria, đều sống sót qua sự kiện K–Pg, mặc dù chịu tổn thất. Đặc biệt, thú có túi gần như biến mất ở Bắc Mỹ, và Deltatheroida châu Á, các họ hàng nguyên thủy của thú có túi còn sống, đã tuyệt chủng. Trong tầng đáy của hệ tầng Hell Creek ở Bắc Mỹ, ít nhất một nửa trong số 10 loài thú răng nhiều mấu đã được ghi nhận và tất cả 11 loài thú có túi không còn được tìm thấy bên trên ranh giới này.
Các loài thú bắt đầu xuất hiện đa dạng vào khoảng 30 triệu năm trước ranh giới K-T, nhưng sự đa dạng của chúng có xu hướng giảm qua thời kỳ này. Một nghiên cứu gần đây cho thấy các loài thú không trải qua sự đa dạng hóa mạnh mẽ qua ranh giới K-T, mặc dù các hốc sinh thái mới được tạo ra bởi sự tuyệt chủng của khủng long. Một số nhóm thú như Chiroptera (dơi) và Cetartiodactyla (bao gồm cá voi, cá heo và động vật guốc chẵn) được cho là đã đa dạng hóa ngay sau ranh giới K-T, nhưng một nghiên cứu mới đây kết luận rằng chỉ có các bộ thú có túi mới thực sự đa dạng hóa sau thời điểm này.
Các loài thú sống quanh thời kỳ K-T thường có kích thước nhỏ, tương đương kích thước của chuột, điều này có thể đã giúp chúng tìm được nơi trú ẩn an toàn. Thêm vào đó, một số loài thú đơn huyệt, thú có túi và thú có nhau thai trước đây có thể đã sống trong môi trường nửa nước nửa cạn hoặc đào hang, vì hiện nay có nhiều dòng thú sinh sống trong các môi trường tương tự. Những loài thú này có thể đã được bảo vệ thêm khỏi áp lực môi trường nghiêm trọng của ranh giới K-T nhờ vào khả năng sống trong các môi trường này.
Bằng chứng
Các hóa thạch ở Bắc Mỹ
Sự kiện tuyệt chủng được phản ánh rõ nét trong các chuỗi trầm tích lục địa Bắc Mỹ qua sự khác biệt đặc trưng giữa các hóa thạch dạng bào phấn từ tầng Maastricht muộn, vốn giàu và phong phú, và sự xuất hiện đột ngột của đỉnh dương xỉ sau ranh giới này.
Chuỗi trầm tích chứa hóa thạch khủng long cung cấp nhiều thông tin nhất về ranh giới K-T trên thế giới nằm ở Bắc Mỹ, đặc biệt là các hệ tầng Hell Creek có tuổi Maastricht muộn ở Montana, Hoa Kỳ. So với các hệ tầng cổ hơn như hệ tầng sông Judith/công viên khủng long (Montana và Alberta, Canada), hệ tầng này cho thấy sự biến đổi trong các quần thể khủng long trong 10 triệu năm cuối của kỷ Phấn Trắng. Các lớp đá hóa thạch này chỉ phân bố một phần ở lục địa.
Các hệ tầng Champagne giữa-muộn chứng minh sự đa dạng cao hơn của các loài khủng long so với các nhóm đá khác. Lớp Maastricht muộn chứa nhiều thành viên của các nhánh chính như: Tyrannosaurus, Ankylosaurus, Pachycephalosaurus, Triceratops và Torosaurus, cho thấy sự phong phú của thức ăn ngay trước khi các loài khủng long tuyệt chủng.
Ngoài các hóa thạch khủng long phong phú, còn có các hóa thạch thực vật cho thấy sự suy giảm về số lượng loài thực vật qua ranh giới K-T. Dưới ranh giới K-T, các hạt phấn hoa của thực vật hạt kín là chủ yếu, nhưng lớp ranh giới chứa ít bào tử phấn hoa và chủ yếu là bào tử dương xỉ. Sự phục hồi của bào tử phấn hoa dần dần diễn ra trên lớp ranh giới. Điều này giống như hiện tượng ở những khu vực bị tàn phá bởi các vụ phun trào núi lửa, nơi sự hồi phục do các loài dương xỉ dẫn đầu trước khi bị thay thế bởi các loài thực vật hạt kín lớn hơn.
Hóa thạch biển
Sự tuyệt chủng hàng loạt của các loài phiêu sinh vật biển có thể đã xảy ra đột ngột ngay tại ranh giới K-T. Một số chi cúc đá đã tuyệt chủng tại hoặc gần ranh giới này; tuy nhiên, một số loài cúc đá cũng bị tuyệt chủng từ từ trước ranh giới K-T do sự thoái lui của biển vào cuối kỷ Creta. Sự giảm dần của các loài hai mảnh vỏ họ Inoceramidae bắt đầu từ trước ranh giới K-T, và sự giảm sút nhỏ, từ từ trong đa dạng loài cúc đá diễn ra trong suốt thời kỳ Creta muộn. Các quá trình này diễn ra đồng thời và kết thúc bằng sự tuyệt chủng hàng loạt đột ngột.
Thời gian xảy ra
Thời điểm xảy ra sự kiện tuyệt chủng vẫn đang gây tranh cãi với hai giả thuyết chính: một giả thuyết cho rằng sự kiện này diễn ra nhanh chóng trong một khoảng thời gian ngắn (vài năm hoặc vài nghìn năm), trong khi giả thuyết khác cho rằng thời gian diễn ra kéo dài hơn. Vấn đề này khó xác định do hiệu ứng Signor-Lipps, cho thấy rằng các di chỉ hóa thạch không hoàn chỉnh và thời điểm tuyệt chủng thực sự của một loài thường xảy ra muộn hơn nhiều so với niên đại của hóa thạch cuối cùng của loài đó. Các nhà khoa học cũng đã phát hiện rất ít lớp đáy liên tục chứa hóa thạch trải qua khoảng thời gian từ vài triệu năm trước đến vài triệu năm sau ranh giới sự kiện tuyệt chủng K–Pg.
Nguyên nhân tuyệt chủng
Nhiều giả thuyết được đưa ra về nguyên nhân gây ra sự tuyệt chủng hàng loạt tại ranh giới K-T, tập trung vào các sự kiện va chạm hoặc hoạt động núi lửa gia tăng, hoặc cả hai. Dựa trên các bằng chứng nghiên cứu, các nhà cổ sinh vật học đồng thuận rằng nguyên nhân chính gây ra sự tuyệt chủng là do một tiểu hành tinh đã va chạm vào Trái Đất cách đây khoảng 65,5 triệu năm, dẫn đến sự tàn phá nghiêm trọng đối với sinh quyển của hành tinh.
Va chạm của tiểu hành tinh
Vào năm 1980, nhóm nghiên cứu do nhà vật lý đoạt giải Nobel Luis Walter Alvarez (con trai của nhà địa chất Walter Alvarez) cùng các nhà hóa học Frank Asaro và Helen Michel đã phát hiện các lớp trầm tích ở ranh giới K-T trên toàn thế giới chứa hàm lượng iridi cao gấp nhiều lần bình thường (từ 20 đến 160 lần trong ba mặt cắt nghiên cứu). Iridi, một nguyên tố hiếm trong vỏ Trái Đất do có xu hướng chìm vào lõi Trái Đất, còn tồn tại nhiều trong các tiểu hành tinh và sao chổi. Do đó, nhóm Alvarez suy đoán rằng một tiểu hành tinh đã va chạm với Trái Đất tại ranh giới K-T. Đây là bằng chứng đầu tiên xác nhận sự kiện va chạm, dù trước đó đã có những suy đoán về khả năng này.
Va chạm này có thể đã làm gián đoạn quá trình quang hợp bằng cách tạo ra đám mây bụi che khuất ánh sáng Mặt Trời và giải phóng các sol khí axit sulfuric vào tầng bình lưu, làm giảm lượng ánh sáng Mặt Trời đến bề mặt Trái Đất từ 10-20%. Phải mất ít nhất 10 năm để các sol khí này phân hủy, thời gian đủ để dẫn đến sự tuyệt chủng của thực vật, sinh vật phiêu sinh và các loài phụ thuộc vào chúng, như thú săn mồi và ăn cỏ. Sinh vật nhỏ ăn mảnh vụn thức ăn có thể có cơ hội sống sót. Sự giải phóng vật liệu va chạm có thể gây ra những cơn bão lửa toàn cầu, với hàm lượng O₂ cao vào cuối kỷ Phấn Trắng giúp hình thành các trận bão này. Hàm lượng O₂ giảm mạnh vào đầu kỷ Tân Cận và các trận bão lửa có thể làm gia tăng CO₂, gây hiệu ứng nhà kính tạm thời sau khi mây bụi lắng đọng, tiêu diệt những sinh vật dễ tổn thương còn sống sót.
Va chạm có thể gây ra mưa axít, tùy thuộc vào loại đá của tiểu hành tinh. Tuy nhiên, nghiên cứu gần đây cho rằng ảnh hưởng này tương đối nhỏ, kéo dài khoảng 12 năm. Môi trường đã trung hòa tính axit, và sự sống sót của các loài dễ tổn thương với mưa axít (như ếch) cho thấy yếu tố này không quan trọng đối với sự tuyệt chủng. Các giả thuyết va chạm chỉ có thể giải thích sự tuyệt chủng diễn ra nhanh chóng vì các đám mây bụi và sol khí lưu huỳnh chỉ tồn tại trong thời gian ngắn khoảng 10 năm.
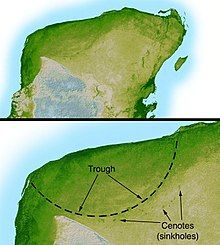
Một nghiên cứu sau này xác nhận hố va chạm Chicxulub dưới bờ biển Yucatán, México, khớp với tuổi giả thuyết của Alvarez. Hố này được phát hiện vào năm 1990 dựa trên nghiên cứu của Glen Penfield từ năm 1978, có hình oval với đường kính khoảng 180 km, gần với kích thước tính toán của nhóm Alvarez. Hình dạng và vị trí của hố cho thấy sự phá hủy sâu xa ngoài đám mây bụi. Tiểu hành tinh va vào đại dương, có thể gây ra siêu sóng thần, dấu vết được tìm thấy ở vùng Caribbe và đông Hoa Kỳ lúc đó là đất liền, và mảnh vụn thực vật, đá lục địa trong trầm tích biển được định tuổi cùng thời với sự va chạm. Thiên thạch va vào lớp thạch cao tạo ra nhiều sol khí lưu huỳnh, làm giảm ánh sáng Mặt Trời và gây mưa axít, tiêu diệt thực vật, phiêu sinh vật và các sinh vật có vỏ calci cacbonat. Vào tháng 2 năm 2008, nhóm nghiên cứu xác định rằng va chạm xảy ra dưới nước ở độ sâu lớn hơn so với giả định trước đây, gia tăng lượng sol khí sulfat và gây ra sự tuyệt chủng hàng loạt do thay đổi môi trường và mưa axít.
Hiện nay, phần lớn các nhà cổ sinh vật học đồng thuận rằng một tiểu hành tinh đã va chạm vào Trái Đất khoảng 65 triệu năm trước. Tuy nhiên, vẫn còn tranh cãi về việc liệu va chạm này có phải là nguyên nhân duy nhất dẫn đến sự tuyệt chủng hàng loạt hay không. Có bằng chứng cho thấy có khoảng 300.000 năm giữa thời điểm va chạm và sự kiện tuyệt chủng hàng loạt. Vào năm 1997, nhà cổ sinh vật học Sankar Chatterjee đã đề xuất sự tồn tại của một hố va chạm lớn hơn có tên Shiva với đường kính 600 km xảy ra đồng thời.
Năm 2007, một giả thuyết mới đã được đưa ra để giải thích sự va chạm tiêu diệt khủng long cách đây 65 triệu năm, liên quan đến nhóm tiểu hành tinh Baptistina. Giả thuyết này đã thu hút sự chú ý từ giới nghiên cứu, một phần vì rất ít mẫu vật của nhóm tiểu hành tinh này được quan sát. Tuy nhiên, một nghiên cứu gần đây chỉ ra rằng 298 Baptistina không có dữ liệu hóa học giống với nguồn gây ra va chạm. Mặc dù việc liên hệ giữa Baptistina và va chạm K-Pg khó xác minh, nhưng cũng không thể loại trừ khả năng đó. Nghiên cứu WISE năm 2011 cho thấy ánh sáng phản xạ từ các tiểu hành tinh này ước tính khoảng 80 triệu năm, không đủ thời gian để thay đổi quỹ đạo và va vào Trái Đất cách đây 65 triệu năm.
Vào tháng 3 năm 2010, một hội thảo quốc tế đã được tổ chức để xem xét các giả thuyết liên quan đến tiểu hành tinh, đặc biệt là va chạm Chicxulub, như nguyên nhân gây ra sự tuyệt chủng. Nhóm 41 nhà khoa học đã xem xét tài liệu trong vòng 20 năm và loại trừ các giả thuyết khác như siêu núi lửa. Họ xác định rằng một khối đá có đường kính 10–15 km đã va chạm với Trái Đất tại Chicxulub, bán đảo Yucatán, México. Va chạm này có thể đã giải phóng năng lượng tương đương với 100 × 10 tấn TNT, lớn hơn khoảng 5 tỷ lần so với năng lượng của bom nguyên tử thả xuống Nagasaki và Hiroshima.
Phun trào núi lửa tại trap Deccan
Trước năm 2000, các lý thuyết cho rằng các dòng lũ bazan trap Deccan đã gây ra sự tuyệt chủng thường liên quan đến quan điểm rằng sự tuyệt chủng diễn ra từ từ, vì các dòng lũ bazan bắt đầu từ khoảng 68 triệu năm trước và kéo dài hơn 2 triệu năm. Tuy nhiên, bằng chứng gần đây cho thấy các trap này đã phun trào kéo dài hơn 800.000 năm qua ranh giới K-T, và có thể đã đóng vai trò trong sự tuyệt chủng cũng như sự hồi phục sinh học chậm sau đó.
Các trap Deccan có thể đã góp phần gây ra sự tuyệt chủng thông qua các cơ chế như phát tán bụi và sol khí sulfuric vào bầu khí quyển, làm giảm lượng ánh sáng Mặt Trời và ảnh hưởng tiêu cực đến quá trình quang hợp của thực vật. Thêm vào đó, hoạt động núi lửa tại trap Deccan có thể đã giải phóng một lượng lớn cacbon dioxide, gây ra hiệu ứng nhà kính khi bụi và sol khí phủ đầy bầu khí quyển.
Khi giả thuyết trap Deccan liên quan đến sự tuyệt chủng từ từ, Luis Walter Alvarez (1911-1988) cho rằng các nhà cổ sinh vật học có thể đã thiếu dữ liệu cần thiết. Dù ban đầu không được chấp nhận rộng rãi, các nghiên cứu sau đó về các lớp đáy hóa thạch đã dần ủng hộ tuyên bố của ông. Đến cuối cùng, phần lớn các nhà cổ sinh vật học đồng thuận rằng sự tuyệt chủng hàng loạt cuối kỷ Phấn Trắng phần lớn hoặc hoàn toàn là do va chạm của tiểu hành tinh vào Trái Đất. Tuy nhiên, Alvarez cũng thừa nhận rằng các yếu tố khác như mực nước biển giảm và sự phun trào núi lửa mạnh mẽ tạo ra các trap Deccan ở Ấn Độ có thể đã góp phần vào sự tuyệt chủng.
Sự va chạm của các tiểu hành tinh nhiều lần
Vào thời điểm ranh giới K-T, có nhiều hố va chạm khác được hình thành. Giả thuyết này cho rằng có thể đã xảy ra nhiều vụ va chạm liên tiếp, có thể là mảnh vỡ của tiểu hành tinh hoặc sao chổi tương tự như sao chổi Shoemaker-Levy 9 đã tạo ra các hố va chạm trên Sao Mộc. Ngoài hố va chạm Chicxulub với đường kính 180 km, còn có hố va chạm Boltysh với đường kính 24 km ở Ukraina (tuổi khoảng 65,17 ± 0,64 triệu năm), hố va chạm Silverpit với đường kính 20 km ở biển Bắc (tuổi khoảng 60–65 triệu năm), và hố va chạm Shiva, có thể có đường kính 600 km, đang còn gây tranh cãi. Những hố va chạm khác có thể đã hình thành trong đại dương Tethys nhưng có thể đã biến mất do các hoạt động kiến tạo như sự di chuyển về phía bắc của mảng châu Phi và Ấn Độ.
Biển thoái Maastricht

Có bằng chứng rõ ràng về sự giảm mực nước biển vào cuối kỷ Phấn Trắng, thấp hơn so với các thời kỳ khác trong Đại Trung sinh. Trong các lớp đá thuộc tầng Maastricht trên toàn cầu, lớp đá phía trên có nguồn gốc lục địa, lớp đá dưới có nguồn gốc ven bờ, và các lớp đá trước đó thuộc đáy biển. Các lớp này không có dấu hiệu nghiêng hay biến dạng kiến tạo, điều này cho thấy sự giảm mực nước biển là nguyên nhân chính. Mặc dù không có bằng chứng trực tiếp về nguyên nhân cụ thể của biển thoái, nhưng giả thuyết hiện tại cho rằng sự giảm hoạt động của các sống núi giữa đại dương có thể là nguyên nhân chính gây ra hiện tượng này.
Sự giảm mực nước biển mạnh có thể đã làm thu hẹp diện tích thềm lục địa, nơi sinh sống của nhiều loài sinh vật biển, góp phần vào sự tuyệt chủng hàng loạt dưới nước. Tuy nhiên, một nghiên cứu khác cho rằng sự thay đổi này có thể không đủ để gây ra sự kiện tuyệt chủng nếu chỉ dựa vào mức độ tuyệt chủng của cúc đá. Sự giảm mực nước biển cũng có thể đã dẫn đến biến đổi khí hậu, bằng cách thay đổi các dòng hải lưu và gió, đồng thời giảm suất phản chiếu của Trái Đất và làm tăng nhiệt độ toàn cầu.
Sự giảm mực nước biển cũng dẫn đến mất các biển nông nằm trong lục địa như Western Interior Seaway ở Bắc Mỹ. Việc này đã gây ra những thay đổi lớn trong môi trường sống, phá hủy các đồng bằng ven biển khoảng 10 triệu năm trước khi sự đa dạng hóa loài xảy ra tại đây, như được ghi nhận trong hệ tầng Dinosaur Park. Hơn nữa, sự mất đi này đã mở rộng các môi trường nước ngọt, vì các dòng chảy trên lục địa phải di chuyển xa hơn trước khi đổ ra biển, tạo điều kiện lý tưởng cho sự phát triển của các loài động vật có xương sống nước ngọt.
Nhiều nguyên nhân khác
J. David Archibald và David E. Fastovsky đã phân tích một kịch bản với ba nguyên nhân chính: hoạt động núi lửa, sự thoái hóa biển, và va chạm của các thiên thạch. Kịch bản này cho rằng các quần thể sinh vật trên cạn và dưới biển đã phải đối mặt với áp lực từ những thay đổi về số lượng và mất môi trường sống. Khủng long, những động vật có xương sống lớn nhất, là nhóm đầu tiên chịu ảnh hưởng khi môi trường thay đổi, dẫn đến sự suy giảm đa dạng của chúng. Đồng thời, bụi từ hoạt động núi lửa làm cho khí hậu ở một số khu vực trở nên lạnh và khô. Cuối cùng, một cú va chạm làm phá vỡ chuỗi thức ăn dựa trên quang hợp cả trên cạn và dưới biển. Giả thuyết này khác biệt với các giả thuyết về từng nguyên nhân riêng lẻ vì nó cho rằng không có nguyên nhân đơn lẻ nào đủ mạnh để gây ra sự tuyệt chủng lớn hay kiểu tuyệt chủng phân loại loài.
- Danh sách năm vụ đại tuyệt chủng
Chú giải
Chú thích
- Tài liệu tham khảo
- Fortey, R (2005). Trái Đất: Một Lịch Sử Gần Gũi. Vintage. ISBN 978-0-375-70620-2.
Các liên kết bên ngoài
 Cổng thông tin về Thiên nhiên
Cổng thông tin về Thiên nhiên Cổng thông tin về Khủng long
Cổng thông tin về Khủng long Trang thông tin về Trái Đất
Trang thông tin về Trái Đất- “Cuộc tranh luận lớn về Chicxulub 2004”. Hội Địa chất London. 2004. Truy cập ngày 2 tháng 8 năm 2007.
- Kring DA (2005). “Sự kiện va chạm Chicxulub: Hiểu biết về ranh giới K–T”. Trung tâm Hình ảnh Không gian NASA. Bản gốc lưu trữ ngày 29 tháng 6 năm 2007. Truy cập ngày 2 tháng 8 năm 2007.
- Cowen R (2000). “Sự tuyệt chủng K–T”. Bảo tàng Sinh học Cổ đại Đại học California. Truy cập ngày 2 tháng 8 năm 2007.
- “Điều gì đã giết chết loài khủng long?”. Bảo tàng Sinh học Cổ đại Đại học California. 1995. Truy cập ngày 2 tháng 8 năm 2007.
| Tiêu đề chuẩn |
|
|---|
