
| Cổ khuẩn | |
|---|---|
| Thời điểm hóa thạch: Đại Tiền Thái cổ - nay 3400–0 Ma Had'n
Archean
Proterozoic
Pha.
| |
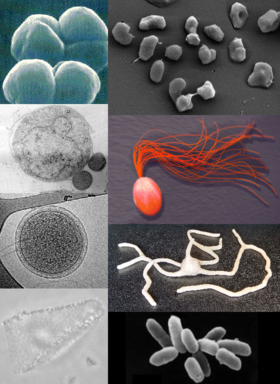
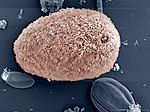
Sự đa dạng của các loài Archaea. | |
| Phân loại khoa học | |
| Liên vực (superdomain) | Prokaryote |
| Vực (domain) | Archaea Woese, Kandler & Wheelis, 1990 |
| Phân nhóm (3 Giới) | |
| |
Vi khuẩn cổ hoặc vi sinh vật cổ (danh pháp khoa học: Archaea) là một nhóm vi sinh vật đơn bào thuộc loại nhân sơ. Chúng thiếu nhân tế bào và các bào quan khác. Trước đây, chúng được coi là một nhóm vi khuẩn đặc biệt và gọi là vi khuẩn cổ (danh pháp khoa học: Archaebacteria), nhưng do có lịch sử tiến hóa độc lập và sự khác biệt sinh hóa so với các dạng sống khác, chúng hiện được phân thành một vực (domain) riêng trong hệ thống ba vực. Ba vực này bao gồm Vi khuẩn (Bacteria), Vi khuẩn cổ và Sinh vật nhân chuẩn (Eukarya), tất cả đều phát sinh từ nguồn gốc chung. Vi khuẩn cổ được chia thành bốn ngành chính, mặc dù có thể còn nhiều hơn. Crenarchaeota và Euryarchaeota là các nhóm được nghiên cứu nhiều nhất. Việc phân loại vi khuẩn cổ vẫn gặp khó khăn vì nhiều loài chưa được nghiên cứu trong phòng thí nghiệm và chỉ mới được phát hiện qua phân tích acid nucleic từ môi trường. Sự phân loại nhóm này với vi khuẩn đã trở nên lỗi thời.
Vi khuẩn cổ và vi khuẩn có nhiều điểm tương đồng về hình dạng và kích thước, mặc dù một số vi khuẩn cổ có hình dáng kỳ lạ như tế bào phẳng hoặc vuông của Haloquadra walsbyi. Mặc dù có vẻ ngoài tương tự vi khuẩn, vi khuẩn cổ có bộ gen và các con đường trao đổi chất tương tự sinh vật nhân chuẩn hơn, đặc biệt là các enzyme liên quan đến phiên mã và dịch mã. Các đặc tính sinh hóa của vi khuẩn cổ rất độc đáo, chẳng hạn như vai trò của các ether lipid trong màng tế bào của chúng. Vi khuẩn cổ có khả năng khai thác nhiều nguồn năng lượng hơn so với sinh vật nhân chuẩn: từ hợp chất hữu cơ như đường đến amonia, ion kim loại và khí hydro. Một số vi khuẩn cổ ưa muối (Halobacteria) sử dụng ánh sáng mặt trời làm nguồn năng lượng, và một số khác có thể cố định cacbon, nhưng không có loài nào thực hiện cả hai chức năng này. Vi khuẩn cổ sinh sản vô tính qua phân đôi, phân mảnh hoặc nảy chồi, khác với vi khuẩn và sinh vật nhân chuẩn, chúng không có bào tử.
Ban đầu, vi khuẩn cổ được biết đến với khả năng sống trong môi trường khắc nghiệt như suối nước nóng và hồ mặn, nhưng sau đó chúng cũng được phát hiện ở nhiều môi trường khác, bao gồm đất, vùng lầy và đặc biệt là đại dương. Trong cộng đồng sinh vật phù du, vi khuẩn cổ có thể là một trong những nhóm sinh vật đông đảo nhất trên Trái Đất. Chúng hiện được xem là một phần quan trọng của sự sống trên hành tinh và có thể đóng vai trò trong chu trình cacbon và nitơ. Mặc dù chưa có ví dụ rõ ràng về vi khuẩn cổ gây bệnh hoặc ký sinh, chúng thường là những sinh vật hỗ sinh hoặc hội sinh. Ví dụ, các loài sinh methan trong ruột người và động vật nhai lại giúp tiêu hóa. Vi khuẩn cổ sinh methan thường được ứng dụng trong sản xuất biogas và xử lý nước thải, và enzyme từ vi khuẩn cổ sống trong điều kiện khắc nghiệt được sử dụng trong công nghệ sinh học.
Phân loại
Vực mới phát hiện

Vào đầu thế kỷ 20, các sinh vật nhân sơ được nhận diện là một nhóm riêng biệt và phân loại dựa trên đặc điểm sinh hóa, hình thái và trao đổi chất. Các nhà vi sinh học đã nỗ lực phân loại các vi sinh vật dựa trên cấu trúc thành tế bào, hình dạng và hợp chất tiêu thụ. Tuy nhiên, vào năm 1965, một phương pháp mới dựa trên trình tự gen đã được đề xuất để xác định mối liên hệ giữa các sinh vật nhân sơ, gọi là phát sinh chủng loại (phylogenetics), và vẫn được sử dụng phổ biến ngày nay.
Vào năm 1977, Carl Woese và George E. Fox đã phân loại Archaea thành một nhóm riêng biệt trong sinh vật nhân sơ, dựa trên trình tự gen của RNA ribosome (rRNA). Ban đầu, hai nhóm được gọi là Vi khuẩn cổ (Archaebacteria) và vi khuẩn thực (Eubacteria), và được phân loại thành các giới (Kingdom) hoặc phân giới (Subkingdom), được Woese và Fox gọi là Urkingdoms. Woese coi nhóm này là một phần cơ bản của sự sống. Để nhấn mạnh sự khác biệt, hai nhóm sau đó được đổi tên thành Archaea và Bacteria. Từ archaea có nguồn gốc từ tiếng Hy Lạp cổ ἀρχαῖα, có nghĩa là 'cổ xưa'.
Ban đầu, chỉ có các vi khuẩn cổ sinh methan được phân loại vào vực mới này, và vi khuẩn cổ được xem như chỉ tồn tại ở môi trường khắc nghiệt như suối nước nóng hoặc hồ mặn. Tuy nhiên, vào cuối thế kỷ 20, các nhà vi sinh học đã nhận ra rằng vi khuẩn cổ là một nhóm sinh vật rộng lớn, đa dạng và phổ biến rộng rãi trong tự nhiên, bao gồm cả môi trường ít khắc nghiệt như đất và đại dương. Sự nhận thức về tầm quan trọng và sự phổ biến của Archaea đến từ việc sử dụng phản ứng chuỗi trùng hợp để phát hiện sinh vật nhân sơ trong các mẫu nước hoặc đất với axít nucleic đơn lẻ, cho phép khám phá và nhận dạng các sinh vật không thể nuôi cấy trong phòng thí nghiệm, điều này vẫn còn thách thức.
Hiện tại về phân loại

Việc phân loại vi khuẩn cổ và sinh vật nhân sơ nói chung đang là một lĩnh vực liên tục thay đổi và có nhiều tranh cãi. Hệ thống phân loại hiện tại chia Archaea thành các nhóm dựa trên đặc điểm cấu trúc và tổ tiên chung. Phân loại này chủ yếu dựa vào kết quả trình tự rRNA để làm sáng tỏ mối quan hệ giữa các sinh vật (phát sinh chủng loại phân tử). Đa số các loài dễ nuôi cấy và nghiên cứu thuộc hai ngành chính là Euryarchaeota và Crenarchaeota. Các nhóm khác mới chỉ được tạo ra tạm thời. Ví dụ, loài Nanoarchaeum equitans, phát hiện năm 2003, đã được xếp vào ngành riêng của nó, Nanoarchaeota. Một ngành mới, Korarchaeota, cũng được đề xuất, gồm các loài ưa nóng đặc biệt có những đặc điểm tương tự hai ngành chính nhưng có mối quan hệ gần hơn với Crenarchaeota. Một số loài mới phát hiện khác như vi sinh vật nano ARMAN, được tìm thấy năm 2006, có mối quan hệ xa hơn với các ngành trên và là một trong những sinh vật nhỏ nhất được biết đến.
Phân loại theo loài
Việc phân loại vi khuẩn cổ thành các loài vẫn còn là vấn đề tranh cãi. Trong sinh học, một loài được định nghĩa là một nhóm cá thể có mối liên hệ gần gũi với nhau. Tiêu chí 'giao phối' thông thường (các cá thể có thể giao phối để tạo ra thế hệ con nhưng không giao phối với các cá thể khác) không áp dụng được cho vi khuẩn cổ vì chúng sinh sản vô tính.
Vi khuẩn cổ cho thấy sự chuyển gen ngang đáng kể giữa các thế hệ. Một số nhà khoa học cho rằng các cá thể có thể tập hợp thành các quần thể tương tự như loài, hình thành bộ gen đồng nhất và có sự chuyển gen không thường xuyên với các tế bào có bộ gen khác biệt, như ở chi Ferroplasma. Ngược lại, nghiên cứu về Halorubrum chỉ ra sự chuyển gen đáng kể với các quần thể ít liên quan, điều này hạn chế việc áp dụng tiêu chí này. Một vấn đề khác cần quan tâm là tầm quan trọng thực tế của việc phân loại loài này.
Những hiểu biết hiện tại về sự đa dạng di truyền còn rời rạc và chưa thể ước lượng chính xác tổng số loài vi khuẩn cổ. Ước tính có từ 18 đến 23 ngành, trong đó chỉ 8 ngành đã có đại diện được nuôi cấy và nghiên cứu. Nhiều nhóm chỉ được xác định qua trình tự rRNA đơn lẻ, cho thấy sự đa dạng của vi khuẩn cổ vẫn còn là một câu hỏi chưa có lời đáp. Các nhóm vi khuẩn cũng chứa nhiều vi sinh vật chưa được nuôi cấy với những đặc tính tương tự, có thể xuất phát từ một nguồn gốc không gian rơi xuống Trái Đất trong thời kỳ sơ khai.
Nguyên gốc và tiến hóa
Mặc dù có các hóa thạch tế bào sinh vật nhân sơ có niên đại lên đến 3,5 tỷ năm trước, hầu hết các sinh vật nhân sơ không có đặc điểm hình thái đặc trưng và hình dạng hóa thạch không thể dùng để xác định chúng là Archaea. Thay vào đó, hóa thạch hóa học của các lipid đặc trưng mang ý nghĩa quan trọng vì những hợp chất này không xuất hiện ở sinh vật khác. Một số tài liệu cho rằng lipid của vi khuẩn cổ hay sinh vật nhân chuẩn đã được phát hiện trong các đá phiến sét có niên đại khoảng 2,7 tỷ năm trước; tuy nhiên, các dữ liệu này vẫn còn nghi vấn. Một số lipid đã được phát hiện trong các mẫu vật từ thời Tiền Cambri. Dấu hiệu cổ nhất được tìm thấy ở vùng Isua, Tây Greenland, nơi lưu giữ các trầm tích cổ xưa nhất trên Trái Đất, hình thành cách đây khoảng 3,8 tỷ năm. Archaea có thể là thế hệ sơ khai nhất xuất hiện trên hành tinh này.
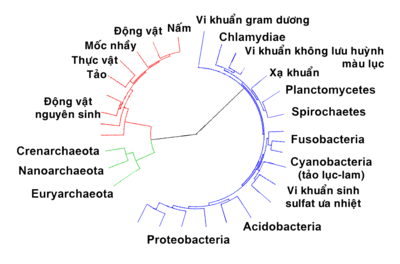
Woese đã chỉ ra rằng vi khuẩn, vi khuẩn cổ và sinh vật nhân chuẩn đều là những dòng tổ tiên khác biệt, phân nhánh từ một nhóm tổ tiên sơ khai. Ngược lại, một số nhà sinh học cho rằng vi khuẩn cổ và sinh vật nhân chuẩn có thể bắt nguồn từ một nhóm vi khuẩn chung. Có khả năng tổ tiên chung cuối cùng của vi khuẩn và vi khuẩn cổ là một sinh vật ưa nhiệt, cho thấy rằng vi khuẩn cổ có thể đã sống trong môi trường nhiệt độ thấp hơn, trong khi các cá thể sống ở môi trường lạnh chỉ xuất hiện sau đó. Khi thấy rằng mối quan hệ giữa vi khuẩn cổ và vi khuẩn còn ít gần gũi hơn so với giữa chúng với sinh vật nhân chuẩn, thuật ngữ prokaryote (sinh vật nhân sơ) chỉ còn có nghĩa là 'không phải nhân chuẩn', làm giảm ý nghĩa của từ này.
Vi khuẩn cổ và sinh vật nhân chuẩn
Mối liên hệ giữa vi khuẩn cổ và sinh vật nhân chuẩn vẫn là vấn đề đang được tranh luận. Mặc dù có những điểm tương đồng về cấu trúc tế bào và chức năng như được trình bày dưới đây, nhiều cây di truyền vẫn phân biệt rõ ràng hai nhóm này.
Các nhận định cho rằng mối quan hệ giữa sinh vật nhân chuẩn và ngành vi khuẩn cổ Euryarchaeota gần gũi hơn mối quan hệ giữa hai ngành Euryarchaeota và Crenarchaeota; và sự hiện diện của các gen giống vi khuẩn cổ trong các loài chắc chắn là vi khuẩn, như Thermotoga maritima, do sự chuyển gen ngang; tất cả tạo ra sự phức tạp cho vấn đề. Một giả thuyết chính cho rằng tổ tiên của sinh vật nhân chuẩn tách ra sớm từ vi khuẩn cổ, và sinh vật nhân chuẩn là kết quả của sự kết hợp giữa một vi khuẩn cổ và một vi khuẩn thực, hình thành nhân và tế bào chất; điều này giải thích nhiều sự tương đồng về di truyền nhưng gặp khó khăn trong việc giải thích cấu trúc tế bào.
Hình thái
Các cá thể vi khuẩn cổ có kích thước đường kính từ 0,1 micromet (μm) đến hơn 15 μm, tồn tại trong nhiều hình dạng như hình cầu, hình que, hình xoắn và hình đĩa. Trong ngành Crenarchaeota, còn có các hình thái đặc biệt như các tế bào thùy với hình dạng bất thường như Sulfolobus, tơ hình kim có đường kính nhỏ hơn nửa micromet như Thermofilum, và tế bào hình que chữ nhật gần như hoàn hảo như Thermoproteus và Pyrobaculum. Haloquadra walsbyi là vi khuẩn cổ có hình dạng vuông và phẳng sống trong môi trường nước mặn cao. Những hình dạng không bình thường này có thể được duy trì nhờ vào thành tế bào và bộ khung tế bào. Những protein liên quan đến bộ khung tế bào của sinh vật khác cũng xuất hiện ở vi khuẩn cổ, với cấu trúc dạng sợi bên trong tế bào, nhưng cấu trúc tế bào của vi khuẩn cổ vẫn còn ít được hiểu biết. Ở Thermoplasma và Ferroplasma, sự thiếu vắng thành tế bào khiến chúng không có hình dạng cố định, và có thể thay đổi hình dạng giống như trùng amip.
Một số loài có khả năng tập hợp hoặc tạo thành các thể sợi dài lên đến 200 μm. Những tế bào này có thể nổi bật trong các biofilm. Đặc biệt, tế bào của Thermococcus coalescens có thể kết hợp với nhau trong quá trình nuôi cấy, hình thành những tế bào khổng lồ. Các cá thể thuộc chi Pyrodictium tạo thành các tập đoàn đa bào phức tạp với hệ thống các ống dài, mỏng và rỗng gọi là cannulae, nằm ngoài bề mặt tế bào và kết nối chúng tạo thành một khối rậm rạp. Mặc dù chức năng của các cannulae vẫn chưa được xác định rõ, chúng có thể hỗ trợ trong việc truyền thông tin hoặc trao đổi dinh dưỡng giữa các tế bào gần nhau. Có các tập đoàn đa loài như cộng đồng 'chuỗi ngọc trai' phát hiện năm 2001 ở một đầm lầy ở Đức, nơi các tập đoàn hình tròn màu trắng thuộc ngành Euryarchaeota được phân bố đều dọc theo các sợi mỏng kéo dài lên đến 15 cm; những sợi này do một loài vi khuẩn đặc biệt tạo ra.
Cấu trúc và chức năng
Vi khuẩn cổ và vi khuẩn có cấu trúc tế bào tương tự nhau, nhưng chức năng và thành phần cấu tạo lại khác biệt. Giống như vi khuẩn, vi khuẩn cổ không có hệ thống màng bên trong và các bào quan. Màng tế bào của vi khuẩn cổ thường liên kết với thành tế bào và chúng di chuyển bằng một hoặc nhiều tiên mao. Về cấu trúc, vi khuẩn cổ gần giống với vi khuẩn gram dương nhất. Hầu hết chúng có một lớp màng sinh chất đơn lẻ và thành tế bào, không có vùng ngoại vi bào chất; một ngoại lệ là Ignicoccus, có vùng chất bào lớn đặc biệt với các túi gắn liền với màng tế bào và được bao bọc bởi một lớp màng ngoài.
Màng tế bào
Màng tế bào của vi khuẩn cổ được cấu thành từ các phân tử hoàn toàn khác biệt so với các dạng sự sống khác, cho thấy mối liên hệ xa giữa vi khuẩn cổ với vi khuẩn và sinh vật nhân chuẩn. Màng tế bào của tất cả các cá thể đều bao gồm các phân tử phospholipid, với một phần đầu phân cực hòa tan trong nước (nhóm phosphate) và phần đuôi không phân cực, không hòa tan (đuôi lipid). Trong môi trường nước, các phospholipid tự tổ chức với phần đầu hướng về nước và đuôi quay vào trong. Kết quả là một lớp màng kép phospholipid, gọi là lớp lipid kép.
Các phospholipid của vi khuẩn cổ có bốn điểm khác biệt chính:
- Vi khuẩn và sinh vật nhân chuẩn sử dụng glycerol-ester lipid để tạo màng tế bào, trong khi vi khuẩn cổ dùng glycerol-ether lipid. Sự khác biệt này nằm ở loại liên kết giữa lipid và nhóm glycerol (màu vàng trong hình). Liên kết ester trong glycerol-ester lipid so với liên kết ether trong glycerol-ether lipid, với liên kết ether bền hơn về mặt hóa học. Điều này giúp vi khuẩn cổ sống sót trong môi trường cực đoan như nhiệt độ cao hoặc pH axít/kiềm. Vi khuẩn và sinh vật nhân chuẩn cũng có một số ether lipid, nhưng chúng không phải là phần chủ yếu của màng tế bào.
- Nhóm glycerol của vi khuẩn cổ có cấu trúc lập thể đảo ngược so với nhóm ở các loài khác. Glycerol có hai dạng enantiomer, giống như bàn tay trái và phải, với các enzyme của vi khuẩn cổ phải thích nghi với dạng đặc biệt này. Điều này gợi ý rằng vi khuẩn cổ sử dụng các enzyme khác biệt trong tổng hợp phospholipid so với vi khuẩn và sinh vật nhân chuẩn, phản ánh sự tách biệt sớm của vi khuẩn cổ.
- Đuôi lipid của vi khuẩn cổ có cấu trúc hóa học đặc biệt, chủ yếu dựa trên chuỗi isoprene dài với nhiều nhánh và đôi khi có vòng cyclopropane hoặc cyclohexane, khác biệt so với axít béo thẳng của các sinh vật khác. Các chuỗi nhánh này giúp bảo vệ màng tế bào khỏi hư hại do nhiệt độ cao, một đặc điểm duy nhất của vi khuẩn cổ.
- Ở một số vi khuẩn cổ, lớp lipid kép được thay thế bằng lớp đơn, với hai phân tử phospholipid kết hợp thành một phân tử duy nhất có hai đầu cực. Điều này giúp màng tế bào thêm ổn định và chống chịu tốt hơn trong môi trường khắc nghiệt, như ở Ferroplasma, nơi các lipid đơn giúp loài này sống sót trong điều kiện axít cao.
Thành tế bào và tiên mao
Hầu hết các vi khuẩn cổ, trừ Thermoplasma và Ferroplasma, đều có thành tế bào. Ở phần lớn vi khuẩn cổ, thành tế bào được kết hợp với các protein bề mặt, hình thành lớp S (S-layer). Lớp S là một hệ thống các phân tử protein được tổ chức cố định bao bọc bên ngoài tế bào như lớp áo giáp, bảo vệ tế bào khỏi các tác động hóa học và vật lý, đồng thời ngăn cản các đại phân tử gắn kết với màng tế bào. Khác với vi khuẩn, hầu hết vi khuẩn cổ không chứa peptidoglycan trong thành tế bào của chúng. Tuy nhiên, có ngoại lệ là pseudopeptidoglycan (peptidoglycan giả) xuất hiện ở vi sinh vật sinh methan, nhưng thiếu axít D-amin và axít N-acetylmuramic.
Tiên mao của vi khuẩn cổ hoạt động tương tự như của vi khuẩn, với cuống dài được điều khiển bởi một động cơ quay ở gốc, lấy năng lượng từ gradient proton trong màng tế bào. Tuy nhiên, tiên mao vi khuẩn cổ có sự khác biệt rõ rệt về cấu trúc và phát triển so với tiên mao vi khuẩn. Hai loại tiên mao tiến hóa từ nguồn gốc khác nhau: tiên mao vi khuẩn liên quan đến hệ thống lưu trữ loại III, trong khi tiên mao vi khuẩn cổ phát triển từ nhung mao vi khuẩn loại IV. Khác với tiên mao vi khuẩn, vốn rỗng và di chuyển từ lỗ trung tâm đến đỉnh, tiên mao vi khuẩn cổ được tổng hợp bằng cách gắn thêm các tiểu đơn vị vào gốc.
Trao đổi chất
Vi khuẩn cổ có khả năng trao đổi chất qua nhiều phản ứng hóa học đa dạng và sử dụng nhiều nguồn năng lượng khác nhau. Các phản ứng này được phân loại theo nhóm dinh dưỡng dựa trên nguồn năng lượng và cacbon. Một số loài lấy năng lượng từ các hợp chất vô cơ như lưu huỳnh và amonia (hóa dưỡng vô cơ), chẳng hạn như loài nitrat hóa, loài sinh methan, và loài oxy hóa methan kị khí. Trong các phản ứng này, một hợp chất chuyển electron cho một hợp chất khác (phản ứng oxy hóa-khử), giải phóng năng lượng để duy trì hoạt động của tế bào. Năng lượng giải phóng được chuyển hóa thành ATP thông qua quá trình thẩm thấu hóa học, tương tự như quá trình xảy ra ở ti thể của tế bào nhân chuẩn.
Một số nhóm vi khuẩn cổ sử dụng ánh sáng mặt trời để tạo năng lượng (quang dưỡng), nhưng chúng không thực hiện quá trình quang hợp để tạo oxy. Nhiều con đường chuyển hóa cơ bản xuất hiện ở nhiều dạng sống khác nhau; chẳng hạn, vi khuẩn cổ sử dụng dạng biến đổi của đường phân (con đường Entner–Doudoroff) và chu trình Krebs hoàn toàn hoặc một phần. Những điểm tương đồng giữa vi khuẩn cổ và các sinh vật khác có thể phản ánh nguồn gốc chung trong lịch sử sự sống và hiệu quả cao của những cơ chế này.
| Loại dinh dưỡng | Nguồn năng lượng | Nguồn cacbon | Ví dụ |
|---|---|---|---|
| Quang dưỡng | Ánh sáng mặt trời | Hợp chất hữu cơ | Halobacteria |
| Hóa dưỡng vô cơ | Hợp chất vô cơ | Hợp chất hữu cơ (hóa dị dưỡng) hoặc cố định cacbon (hóa tự dưỡng) | Ferroglobus, Methanobacteria và Pyrolobus |
| Hóa dưỡng hữu cơ | Hợp chất hữu cơ | Hợp chất hữu cơ hoặc cố định cacbon | Pyrococcus, Sulfolobus hoặc Methanosarcinales |
Một số loài Euryarchaeota, như các loài sinh methan, sống trong môi trường kị khí như đầm lầy. Trao đổi chất này đã tiến hóa từ rất sớm và có thể là đặc điểm của những sinh vật sống tự do đầu tiên. Quá trình sinh methan sử dụng khí cacbonic làm chất nhận điện tử để oxy hóa khí hydro, và liên quan đến một loạt coenzyme đặc trưng như coenzyme M và methanofuran. Các hợp chất hữu cơ khác như rượu, acid acetic và acid formic cũng được sử dụng làm chất nhận điện tử. Các phản ứng này cũng xảy ra ở các vi khuẩn cổ trong ruột. Acid acetic có thể bị phân hủy trực tiếp thành methan và cacbon dioxide nhờ các vi khuẩn cổ 'dưỡng acetic' thuộc bộ Methanosarcinales, đóng vai trò chính trong cộng đồng vi sinh vật sản sinh biogas.
Các vi khuẩn cổ khác sử dụng CO
2 trong khí quyển làm nguồn cacbon thông qua quá trình cố định cacbon (tự dưỡng). Quá trình này bao gồm dạng biến đổi của chu trình Calvin hoặc một con đường chuyển hóa mới gọi là chu trình 3-hydroxypropionate/4-hydroxybutyrate. Crenarchaeota sử dụng chu trình Krebs ngược, trong khi Euryarchaeota áp dụng con đường acetyl-CoA thu gọn. Quá trình cố định cacbon sử dụng năng lượng từ nguồn vô cơ. Không có vi khuẩn cổ nào được biết đến với khả năng quang hợp. Các nguồn năng lượng của vi khuẩn cổ rất đa dạng, từ oxy hóa amonia ở Nitrosopumilales đến oxy hóa hydro sulfite hoặc sulfide nguyên tử ở các loài thuộc Sulfolobus, với việc sử dụng cả oxy và ion kim loại làm chất nhận điện tử.
Vi khuẩn cổ quang dưỡng sử dụng ánh sáng để sản xuất năng lượng hóa học dưới dạng ATP. Ở Halobacteria, các chất truyền ion như bacteriorhodopsin và halorhodopsin được kích hoạt bởi ánh sáng để tạo ra gradient ion bằng cách bơm ion ra khỏi tế bào qua màng sinh chất. Năng lượng lưu trữ trong các gradient điện hóa này sau đó được chuyển thành ATP thông qua quá trình tổng hợp ATP, một dạng phosphoryl quang hóa. Khả năng của các bơm ánh sáng này trong việc di chuyển ion qua màng phụ thuộc vào sự biến đổi ánh sáng trong cấu trúc của cofactor retinol trung tâm protein.
Di truyền
Vi khuẩn cổ thường có một nhiễm sắc thể dạng vòng đơn, kích thước có thể lên tới khoảng 5.751.492 cặp base như ở Methanosarcina acetivorans, bộ gen lớn nhất trong nhóm này. Ngược lại, loài Nanoarchaeum equitans, với bộ gen nhỏ nhất, chỉ có 490.885 cặp base và chứa khoảng 537 gen mã hóa protein. Các mảnh DNA nhỏ hơn, gọi là plasmid, cũng xuất hiện ở vi khuẩn cổ. Plasmid có thể di chuyển giữa các tế bào thông qua tương tác vật lý, tương tự như sự tiếp hợp ở vi khuẩn.
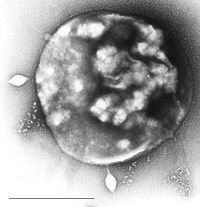
Vi khuẩn cổ có thể bị nhiễm các virus DNA chuỗi kép, có những hình dạng bất thường như dạng chai, que có móc hoặc hình giọt nước và không có liên quan đến các dạng virus khác. Các virus này đã được nghiên cứu chi tiết ở các loài ưa nhiệt, đặc biệt là các bộ Sulfolobales và Thermoproteales. Vào năm 2009, một virus DNA chuỗi đơn xâm nhiễm loài ưa muối đã được phát hiện. Để phòng chống các loại virus này, có thể cần đến can thiệp RNA từ các trình tự DNA lặp có liên quan đến các gen virus.
Vi khuẩn cổ có đặc điểm di truyền rất khác biệt so với vi khuẩn và sinh vật nhân chuẩn, với khoảng 15% protein mã hóa bởi bộ gen vi khuẩn cổ là đặc hữu cho nhóm này, dù vai trò của phần lớn các gen này vẫn chưa được xác định. Trong số các protein độc nhất có chức năng đã được biết đến, hầu hết đều liên quan đến sự sinh methan. Những protein chung giữa vi khuẩn cổ, vi khuẩn và sinh vật nhân chuẩn đều liên quan đến chức năng cơ bản của tế bào, như phiên mã, dịch mã và chuyển hóa nucleotide. Các đặc trưng riêng của vi khuẩn cổ bao gồm tổ chức gen liên quan đến chức năng, các enzyme xúc tác các bước trong cùng con đường chuyển hóa thuộc operon mới, cùng với sự khác biệt rõ rệt trong gen của tRNA và enzyme aminoacyl tRNA synthetase.
Phiên mã và dịch mã ở vi khuẩn cổ tương đồng hơn với sinh vật nhân chuẩn so với vi khuẩn, vì RNA polymerase và ribosome của chúng gần giống với của sinh vật nhân chuẩn. Mặc dù vi khuẩn cổ chỉ có một loại RNA polymerase duy nhất, cấu trúc và chức năng của nó trong phiên mã gần gũi với RNA polymerase II của sinh vật nhân chuẩn, nhờ vào các protein tương tự (nhân tố phiên mã chung) giúp gắn kết RNA polymerase với promoter của gen. Tuy nhiên, các nhân tố phiên mã khác của vi khuẩn cổ lại giống hơn với của vi khuẩn. Sự chỉnh sửa sau phiên mã ở vi khuẩn cổ đơn giản hơn so với sinh vật nhân chuẩn, khi hầu hết các gen của vi khuẩn cổ không có intron, mặc dù intron có thể xuất hiện trong các gen RNA vận chuyển và RNA ribosome, cũng như trong một số gen mã hóa protein.
Sinh sản
Vi khuẩn cổ sinh sản vô tính qua các phương pháp phân đôi, phân mảnh hoặc nảy chồi; không có giảm phân, vì vậy nếu một loài vi khuẩn cổ có nhiều dạng, tất cả đều chia sẻ cùng một bộ gen. Quá trình phân bào được kiểm soát trong chu trình tế bào; sau khi nhiễm sắc thể nhân đôi và tách thành hai nhiễm sắc thể con, tế bào mới sẽ phân chia. Các nghiên cứu mới chỉ được thực hiện trên chi Sulfolobus, nhưng chu trình tế bào ở đây có những đặc điểm tương tự như ở vi khuẩn và sinh vật nhân chuẩn. Các nhiễm sắc thể nhân đôi từ nhiều điểm khởi đầu, sử dụng DNA polymerase giống với enzyme tương tự của sinh vật nhân chuẩn. Tuy nhiên, các protein điều khiển phân chia tế bào, như FtsZ tạo thành vòng co xung quanh tế bào, và các thành phần vách ngăn dựng lên dọc giữa trung tâm tế bào, tương đồng với các protein ở vi khuẩn.
Vi khuẩn cổ không có bào tử như vi khuẩn và sinh vật nhân chuẩn. Một số loài Haloarchaea có thể thay đổi hình thái và phát triển thành các kiểu tế bào khác nhau, bao gồm các cấu trúc vách mỏng giúp chống chịu với áp lực thẩm thấu và tồn tại trong môi trường nước có độ muối thấp, nhưng những cấu trúc này không phải là cấu trúc sinh sản và có thể giúp vi khuẩn cổ di chuyển đến môi trường sống mới.
Sinh thái học
Môi trường sống
Vi khuẩn cổ hiện diện ở nhiều loại môi trường khác nhau và là thành phần quan trọng trong các hệ sinh thái toàn cầu, có thể chiếm gần 20% sinh khối của Trái Đất. Những vi khuẩn cổ đầu tiên được phát hiện sống trong điều kiện cực đoan. Một số loài có khả năng tồn tại ở nhiệt độ rất cao, thường trên 100°C (212°F), và được tìm thấy ở các mạch nước phun, ống khói đen ở miệng thủy nhiệt và giếng dầu. Chúng cũng sống trong các môi trường cực lạnh, có độ mặn, tính acid hoặc tính kiềm cao. Bên cạnh đó, một số loài vi khuẩn cổ sống trong điều kiện bình thường như đất, cống rãnh và đại dương.

Vi khuẩn cổ sống trong môi trường cực đoan được phân chia thành bốn nhóm sinh lý chính: ưa mặn, ưa nhiệt, ưa kiềm và ưa acid. Các nhóm này không hoàn toàn tách biệt và không loại trừ lẫn nhau, vì một số vi khuẩn cổ có thể thuộc nhiều nhóm khác nhau. Dù vậy, chúng là cơ sở hữu ích cho việc phân loại.
Các loài ưa mặn, chẳng hạn như chi Halobacterium, sống trong môi trường có nồng độ muối rất cao như hồ mặn, và chúng thường có số lượng vượt trội so với nhóm vi khuẩn tương đương ở mức muối cao hơn 20–25%. Các loài ưa nhiệt phát triển tốt nhất ở nhiệt độ trên 45°C (113°F), thường thấy ở suối nước nóng; những vi khuẩn cổ 'siêu ưa nhiệt' tối ưu hóa sự phát triển ở nhiệt độ trên 80°C (176°F). Loài vi khuẩn cổ Methanopyrus kandleri có khả năng sống ở nhiệt độ kỷ lục 122°C (252°F), là mức nhiệt độ cao nhất ghi nhận được.
Một số vi khuẩn cổ tồn tại trong môi trường có độ acid hoặc kiềm cao. Ví dụ, loài vi khuẩn cổ ưa acid cực đoan như Picrophilus torridus có thể sống trong điều kiện pH 0, tương đương với sự tồn tại trong acid sulfuric nồng độ 1,2 mol.
Khả năng chống chịu với môi trường khắc nghiệt đã khiến vi khuẩn cổ trở thành đối tượng nghiên cứu quan trọng trong việc tìm kiếm sự sống ngoài Trái Đất. Những môi trường cực đoan mà chúng sống có sự tương đồng với điều kiện trên Sao Hỏa, dẫn đến giả thuyết rằng vi sinh vật có thể di chuyển giữa các hành tinh thông qua các mảnh vỡ không gian.
Gần đây, các nghiên cứu cho thấy vi khuẩn cổ không chỉ sống ở môi trường nhiệt độ cao hay bình thường mà còn phổ biến ở môi trường lạnh. Chúng có mặt, đôi khi với số lượng lớn, trong các vùng đại dương lạnh như biển hai cực. Đặc biệt, vi khuẩn cổ xuất hiện với số lượng lớn (lên tới 40% sinh khối vi sinh) trong các cộng đồng sinh vật phù du (như picoplankton – sinh vật phù du kích cỡ 0,2-2 µm). Tuy nhiên, hầu hết chúng chưa được phân lập và nghiên cứu đầy đủ, dẫn đến hiểu biết hạn chế về vai trò của chúng trong sinh thái đại dương và ảnh hưởng của chúng đối với các chu trình sinh địa hóa toàn cầu. Một số loài Crenarchaeota ở biển có khả năng nitrit hóa, cho thấy chúng có thể ảnh hưởng đến chu trình nitơ đại dương, mặc dù chúng cũng có thể sử dụng nguồn năng lượng khác. Số lượng lớn vi khuẩn cổ được phát hiện trong các trầm tích dưới đáy biển, nơi chúng chiếm ưu thế ở độ sâu hơn 1 mét dưới mặt lớp trầm tích.
Vai trò trong chu trình hóa học
Vi khuẩn cổ đóng vai trò quan trọng trong việc tái sử dụng các nguyên tố như cacbon, nitơ và sulfide qua các môi trường sống đa dạng của chúng. Mặc dù các hoạt động này cần thiết cho chức năng hệ sinh thái bình thường, vi khuẩn cổ cũng có thể ảnh hưởng đến những thay đổi do con người gây ra, bao gồm cả ô nhiễm.
Vi khuẩn cổ tham gia vào nhiều giai đoạn của chu trình nitơ, từ việc tách nitơ ra khỏi hệ sinh thái qua các quá trình như hô hấp dùng nitơ và phản nitrat hóa, đến việc hấp thụ nitơ qua đồng hóa và cố định nitơ. Gần đây, vai trò của vi khuẩn cổ trong các phản ứng oxy hóa amonia đã được phát hiện, đặc biệt là trong đại dương. Chúng cũng đóng vai trò quan trọng trong việc oxy hóa amonia ở đất, sản xuất nitrit mà sau đó được chuyển hóa thành nitrat bởi các vi sinh vật khác. Thực vật và các sinh vật khác sau đó sẽ sử dụng những hợp chất này.
Trong chu trình sulfide, vi khuẩn cổ như Sulfolobus oxy hóa các hợp chất lưu huỳnh để giải phóng nguyên tố này khỏi đá, giúp các loài khác sử dụng. Tuy nhiên, quá trình này cũng sản xuất acid sulfuric như một sản phẩm phụ, và sự phát triển của các loài này ở các mỏ bỏ hoang có thể tạo ra nước acid mỏ, gây hại cho môi trường.
Trong chu trình cacbon, vi khuẩn cổ sinh methan chuyển đổi hydro và đóng vai trò quan trọng trong phân hủy vật chất hữu cơ ở các hệ sinh thái kị khí như trầm tích, đầm lầy và trong xử lý nước thải. Methan là một trong những khí nhà kính chính, chiếm khoảng 18% tổng lượng khí nhà kính toàn cầu, và có hiệu ứng nhà kính mạnh gấp 25 lần so với khí cacbonic. Các loài sinh methan là nguồn chính của khí methan, góp phần lớn vào lượng phát thải methan hàng năm, từ đó ảnh hưởng đến hiện tượng ấm lên toàn cầu.
Tương tác với các loài khác

Quan hệ giữa Archea và các sinh vật khác có thể là hỗ sinh hoặc hội sinh. Đến năm 2007, chưa có bằng chứng nào về vi khuẩn cổ gây bệnh hay ký sinh. Tuy nhiên, đã có giả thuyết rằng một số loài sinh methan có thể liên quan đến các bệnh nhiễm trùng miệng; ví dụ như Nanoarchaeum equitans có thể là ký sinh của một số loài vi khuẩn cổ khác vì nó chỉ sống và sinh sản khi nằm trong tế bào của loài Ignicoccus hospitalis thuộc nhóm Crenarchaeota mà không mang lại lợi ích gì cho vật chủ.
Hỗ sinh
Một ví dụ rõ ràng về hỗ sinh là sự tương tác giữa động vật nguyên sinh và vi khuẩn cổ sinh methan trong hệ tiêu hóa của động vật tiêu hóa cellulose, như động vật nhai lại và mối. Trong môi trường kị khí, các động vật nguyên sinh phá vỡ cellulose thực vật để thu năng lượng, giải phóng khí hydro dư thừa. Khi vi khuẩn sinh methan chuyển hóa khí hydro thành methan, động vật nguyên sinh có thể thu được nhiều năng lượng hơn.
Ở những loài động vật nguyên sinh kị khí như Plagiopyla frontata, vi khuẩn cổ sống bên trong và tiêu thụ khí hydro sản xuất trong hydrogenosome của chúng. Vi khuẩn cổ cũng có thể cộng sinh với các sinh vật lớn hơn; ví dụ, loài vi khuẩn cổ sống ở biển Cenarchaeum symbiosum cộng sinh trong loài bọt biển Axinella mexicana.
Hội sinh
Vi khuẩn cổ có thể tồn tại theo kiểu hội sinh, thu lợi từ sự liên kết mà không ảnh hưởng đến loài khác. Ví dụ, vi khuẩn methan Methanobrevibacter smithii là loài phổ biến nhất trong cơ thể người, chiếm khoảng 10% tổng số vi sinh vật nhân sơ trong ruột. Những vi khuẩn methan này có thể hỗ sinh cho các sinh vật khác trong ruột, hỗ trợ quá trình tiêu hóa. Chúng cũng kết hợp với nhiều sinh vật khác trên san hô và trong đất xung quanh rễ cây.
Vai trò trong công nghiệp và kỹ thuật
Vi khuẩn cổ sống trong môi trường khắc nghiệt, chịu được nhiệt độ cao, acid và kiềm, cung cấp các enzyme hoạt động tốt dưới điều kiện cực đoan. Ví dụ, Pfu DNA polymerase từ Pyrococcus furiosus đã cách mạng hóa sinh học phân tử bằng cách cho phép phản ứng chuỗi trùng hợp trở nên đơn giản và nhanh chóng hơn. Trong công nghiệp, các enzyme như amylase, galactosidase và pullulanase từ các loài Pyrococcus hoạt động ở trên 100°C hỗ trợ sản xuất thực phẩm, như sữa và váng sữa có lượng lactose thấp. Enzyme từ vi khuẩn cổ ưa nhiệt cũng ổn định trong dung môi hữu cơ, có ứng dụng trong hóa học xanh và sinh học cấu trúc.
Mặc dù enzyme từ vi khuẩn cổ có nhiều ứng dụng, việc sử dụng chính các loài vi khuẩn cổ trong công nghệ sinh học vẫn còn hạn chế. Vi khuẩn cổ sinh methane đóng vai trò quan trọng trong xử lý nước thải và sản xuất biogas. Trong khai thác mỏ, vi khuẩn cổ ưa acid có tiềm năng chiết xuất kim loại từ quặng như vàng, cobalt và đồng.
Vi khuẩn cổ sở hữu một tập hợp mới các chất kháng sinh có triển vọng ứng dụng. Một số đã được phát hiện, nhưng còn hàng trăm chất khác chưa được khám phá, đặc biệt trong các nhóm Haloarchaea và Sulfolobus. Những hợp chất này có cấu trúc độc đáo so với kháng sinh từ vi khuẩn, có thể có cơ chế hoạt động mới và giúp phát triển các gen đánh dấu chọn lọc trong sinh học phân tử của vi khuẩn cổ.
Chú giải
Đọc thêm
- Howland, John L. (2000). Những Khám Phá Đáng Ngạc Nhiên về Archaea: Khám Phá Một Miền Sống Mới. Oxford: Oxford University Press. ISBN 0-19-511183-4.
- Martinko JM, Madigan MT (2005). Sinh Học Vi Sinh Vật của Brock. Englewood Cliffs, N.J: Prentice Hall. ISBN 0-13-144329-1.
- Garrett RA, Klenk H (2005). Archaea: Tiến Hoá, Sinh Lý và Sinh Học Phân Tử. WileyBlackwell. ISBN 1-40-514404-1.
- Cavicchioli R (2007). Archaea: Sinh Học Phân Tử và Tế Bào. American Society for Microbiology. ISBN 1-55-581391-7.
- Blum P (biên tập) (2008). Archaea: Các Mô Hình Mới cho Sinh Học Vi Khuẩn. Caister Academic Press. ISBN 978-1-904455-27-1.
- Lipps G (2008). “Plasmid Cổ Khuẩn”. Plasmids: Nghiên Cứu Hiện Tại và Xu Hướng Tương Lai. Caister Academic Press. ISBN 978-1-904455-35-6.
Các liên kết bên ngoài
- Archaea (Prokaryote) trên Encyclopædia Britannica (tiếng Anh)
- Giới thiệu về Archaea, sinh thái, phân loại và cấu trúc
- Cổ khuẩn trên Encyclopedia of Life
- Cổ khuẩn tại Trung tâm Thông tin Công nghệ Sinh học Quốc gia Hoa Kỳ (NCBI)
- Cổ khuẩn 935939 trên Hệ thống Thông tin Phân loại Tích hợp (ITIS)
- Các tên hợp lệ của chi trong miền (hoặc đế) Archaea (hoặc Archaeobacteria) LPSN: Các chi của Archaea
- Tree of Life thể hiện mối quan hệ của Archaea với các dạng sống khác
- Tìm kiếm bất kỳ bộ gen vi khuẩn cổ nào trên UCSC





Eukaryota | |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
Vực: Archaea • Bacteria • Eukaryota | |||||||||||||||||||||||||
| B i k o n t a |
| ||||||||||||||||||||||||
| U n i k o n t a |
| ||||||||||||||||||||||||
| Incertae sedis |
| ||||||||||||||||||||||||

Phân loại vi khuẩn cổ |
|---|


Sự sống hiện hữu trên Trái Đất |
|---|





Các thành phần tự nhiên |
|---|
| Tiêu đề chuẩn |
|
|---|
| Thẻ nhận dạng đơn vị phân loại |
|---|